Portaalsite voor de échte zeeaquariaan
Mesofotische koraalriffen, het leven in de diepte
Mesofotische koraalriffen, het leven in de diepte
Door Tim Wijgerde, Ph. D., Foto’s: Pim Bongaerts Ph. D., tenzij anders vermeld. Vertaling: Germain Leys
Recente wetenschappelijke bevindingen suggereren dat deze zogenaamde mesofotische riffen kunnen fungeren als toevluchtsoord voor veel rifsoorten, waaronder vissen en koralen, die kunnen helpen om ondiepe riffen opnieuw te bevolken na perioden van omgevingsstress. Dit artikel beschrijft enkele van de mesofotische koraalriffen die over de hele wereld worden gevonden, geïllustreerd met foto’s gemaakt tijdens diepe duiken.

Een soortenrijk mesofotisch rif rond het eiland Bunaken, Sulawesi, op een diepte van 40 m (133 ft.)
Hoewel koraalriffen over de hele wereld zijn bestudeerd sinds de tijd van Charles Darwin (1809-1882), heeft het vele jaren geduurd voordat we een basiskennis van deze kwetsbare ecosystemen ontwikkelden. Men is het er nu over eens dat de vele vormen van symbiose tussen koralen, sponzen, zoöxanthellen, bacteriën en andere soorten de sleutel zijn tot het succes van koraalriffen. Door te groeien in het mariene equivalent van een woestijn, met zeer lage concentraties anorganische nutriënten, kunnen door de constante uitwisseling en recycling van materie riffen blijven bestaan (De Goeij et al. 2013).

Een verticale rotswand, zwaar bedekt met sponzen. Let op de gele pincetvis, Forcipiger flavissimus in het midden.
Koraalriffen worden meestal afgeschilderd als kleurrijke ecosystemen die groeien in warme, heldere en ondiepe wateren, en zo genieten we er gewoonlijk van tijdens het duiken of snorkelen. Wat minder bekend is, is dat veel riffen zich in feite uitstrekken tot in diepere wateren, met zoöxanthellaat-koralen die tot een diepte van minstens 165 meter (550 voet) worden gevonden. Deze zogenaamde mesofotische riffen (van de Griekse woorden mésos, of middle, en phot, of licht) worden momenteel gedefinieerd als voorkomend tussen 30 en 200 meter diep (Olson en Kellogg 2010). Ze herbergen talloze dier- en plantensoorten, en sommige komen alleen voor in deze diepere wateren.


Links: Vanwege hun ontoegankelijkheid zijn mesofotische riffen het onderwerp geweest van beperkt wetenschappelijk onderzoek. Tegenwoordig stellen moderne technologieën zoals rebreathers, submersibles en ROV’s wetenschappers in staat om zelfs de diepste riffen te verkennen. In de komende jaren zullen zeebiologen meer ontdekken over de ecologie en biodiversiteit van mesofotische riffen, evenals over de genetica en fysiologie van de belangrijkste leden, de steenkoralen. Hier exploiteert Dr. Pim Bongaerts van het XL Catlin Seaview Survey-team een ROV op het Great Barrier Reef. Foto: Richard Vevers.
Rechts: Duik in de diepte. In de afgrond wacht veel zeeleven op ontdekking. Let op de verschillende zeelelies op de bergkam.
Diepe refugia
Wat mesofotische riffen belangrijk maakt, is hun hoge biodiversiteit, evenals hun potentieel om te dienen als diepe refugia, veilige havens, voor veel soorten. Dit komt omdat hun diepte hen minder vatbaar maakt voor natuurlijke en menselijke invloeden (Bak et al. 2005; Bongaerts et al. 2010, 2011a, b). Er zijn bijvoorbeeld aanwijzingen dat diepe riffen kunnen ontsnappen aan schadelijke stormen en koraalverbleking, hoewel ze niet volledig immuun zijn voor stressfactoren.
Recent bewijs toont aan dat ongeveer 25% van alle zoöxanthellate koraalsoorten, d.w.z. koralen die symbiotische dinoflagellaten van het genus Symbiodinium herbergen, een diepteverdeling heeft van ondiepe wateren tot mesofotische diepten van 30-60 meter (van Oppen et al.2011). In Noordoost-Australië, Muir et al. (2015) ontdekten dat van de 76 Acropora- en Isopora-soorten die in ondiepe wateren zijn geregistreerd, 21% zich uitstrekt tot mesofotische diepten. Deze zogenaamde diepte-generalistische soorten kunnen helpen bij het herstel van het rif door larven en gameten uit de diepte vrij te laten, wat uiteindelijk zou kunnen leiden tot rekrutering van nieuwe koraalkolonies op ondiepe riffen. Inderdaad, met behulp van DNA-analyse, van Oppen et al. (2011) ontdekten dat het broedende koraal Seriatopora hystrix larven vrijgeeft in diepere wateren die zich nestelen op ondiepe riffen.

Een diep Indonesisch rif, dichtbevolkt door steenkoralen, gorgonen, zachte koralen, sponzen, zeelelies, koraalalgen en juffers. Dergelijke riffen kunnen fungeren als een “veilig huis” voor kwetsbare soorten.
Ze ontdekten echter ook dat dit fenomeen afhankelijk is van het onderzochte rif. Het herstel van ondiep rif door koraallarven uit de diepte wordt dus beperkt door het soortaantal (diepte-generalisten) en komt niet noodzakelijk overal voor.
Via de volgende link kun je een video volgen van zulk een duik:
https://www.youtube.com/watch?v=AAVTynS4lIo&feature=player_embedded

Een eenzame majestueuze keizervis Pomacanthus navarchus foerageert op een mesofotisch rif, met grote groepen Pachyseris spp.
Omgeving: licht, temperatuur, waterstroom en voedingsstoffen
Mesofotische riffen strekken zich veel verder uit dan hun ondiepe tegenhangers, en het zijn verschillende werelden. Beneden 30 meter vervagen warme, zonovergoten wateren snel tot een sombere kou. Koralen die op 100 m diepte (333 ft.) En lager groeien, moeten bijvoorbeeld overleven in een omgeving met minder dan 1% van de maximale zonnestraling, temperaturen van 20 graden Celsius (68 ° F) en lager, een beperkte waterstroom en verhoogde voedingsstoffen. Als iemand een donker, onverwarmd aquarium zou opzetten met nauwelijks waterstroom en geen filtratie, en het zou vullen met steenkoralen uit de diepte, kan het een paar wenkbrauwen optrekken. Dit zou echter in de buurt komen van de omstandigheden die koralen uit de diepere mesofotische zone ervaren. Hieronder zal ik vier belangrijke abiotische factoren bespreken die de mesofotische zone bepalen, met een diepte tussen 30 en 200 meter (100-667 voet).

Qua diepte kunnen koraalriffen grofweg worden onderverdeeld in drie hoofdtypen; ondiepe riffen, met een dieptebereik van 0-30 meter (0-100 voet), mesofotische riffen, die voorkomen tussen 30 en 200 meter diep (100-667 voet), en diepe riffen, gevonden onder de 200 meter (667 voet). Ondiepe riffen herbergen een breed scala aan lichtafhankelijke koralen, sponzen en (macro) algen. Mesofotische riffen worden bevolkt door minder koraalsoorten, met vaak een hoger bedekkingspercentage. Hier hebben koralen hun vorm en fysiologie aangepast aan deze slecht verlichte omgeving, met afgeplatte kolonies en een hogere lichtopvangefficiëntie. Zowel ondiepe als mesofotische riffen hebben een combinatie van zoöxanthellate en azooxanthellate koralen. Beneden 200 meter dringt vrijwel geen licht door en dus missen bijna alle koralen die hier worden gevonden zoöxanthellen. In deze diepe wateren is het scleractijnse koraal Lophelia pertusa een belangrijke rifbouwer. Pijlen geven gegeneraliseerde gradiënten aan in termen van lichtintensiteit, temperatuur en waterstroomsnelheid, die afnemen met de diepte, en anorganische voedingsstoffen, die toenemen met de diepte. Afbeelding aangepast van Lesser et al. (2009) en Olson en Kellogg (2010), met foto’s van Wikimedia Commons / Lophelia II 2010 Expedition / NOAA-OER / BOEMRE / Muir et al. (2015).
Licht
Wanneer we de oceaan induiken, wordt zowel de intensiteit (bestraling) als de spectrale kwaliteit van licht beïnvloed. Hoe dieper we gaan, hoe lager de lichtintensiteit, aangezien licht wordt geabsorbeerd en verstrooid door zeewater en zijn bestanddelen. Op een zonnige tropische dag, rond het middaguur, is de lichtintensiteit aan het oppervlak ongeveer 2.500 µmol fotonen m-2 s-1, gemeten als fotosynthetisch actieve straling (of PAR, ~ 400-700 nm, Frade et al. 2008; Lesser et al. 2010). Op een diepte van 5 meter is dit al afgenomen tot ongeveer 1.000 µmol fotonen m-2 s-1, en op een typische mesofotische diepte van 60 meter (200 voet) is de bestralingssterkte aanzienlijk gedaald tot slechts 50 µmol fotonen m- 2 s-1.
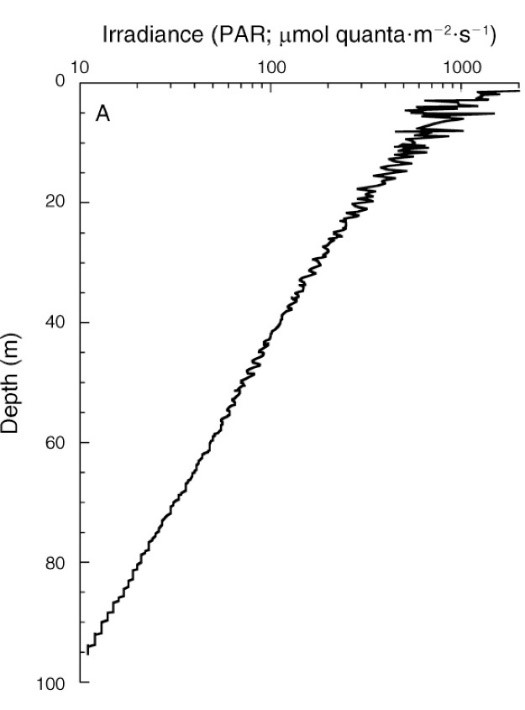
Instralingsdiepteprofiel tussen 0 en 95 m (0-317 ft.) Bij Bock Wall, Lee Stocking Island, Bahama’s. De bestralingssterkte van het oppervlak was 2.478 µmol fotonen m-2 s-1 (~ 400-700 nm). Merk op dat de x-as is uitgezet op een logschaal, wat resulteert in een rechte curve, en dat PAR-niveaus worden uitgedrukt als absolute waarden. Van Lesser et al. (2010).
Om deze waarden in perspectief te plaatsen: de meeste rifaquaria ervaren lichtintensiteiten in het bereik van 100 tot 1.000 µmol fotonen m-2 s-1, wat overeenkomt met waterdiepten van ongeveer 40 tot 5 meter (133 tot 17 voet). Het is echter belangrijk op te merken dat in het veld het bestralingsprofiel parabolisch is, d.w.z. de lichtintensiteit is ‘s ochtends en’ s avonds laag en piekt ‘s middags. Bovendien worden de stralingsniveaus in de natuur aanzienlijk beïnvloed door seizoen, breedtegraad en weer. Op een bewolkte dag dringt er bijvoorbeeld beduidend minder licht door het water, dat ook verschuift naar het blauwe uiteinde van het lichtspectrum. Waterhelderheid heeft ook een grote invloed op de diepteverdeling van koralen.
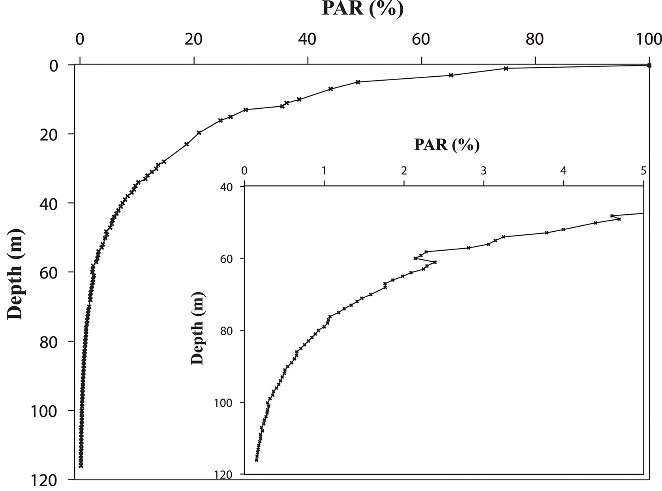
Diepteprofiel van de bestralingssterkte tussen 0 en 115 m (0-383 ft.) In de Golf van Aqaba voor het Interuniversitair Instituut in Eilat in oktober, gemeten als fotosynthetisch actieve straling (PAR, ~ 400-700 nm). De bestralingssterkte van het oppervlak was relatief laag tijdens deze meting vanwege de tijd van het jaar, bij 1.400 µmol fotonen m-2 s-1. Merk op dat de x-as is uitgezet op een lineaire schaal, wat resulteert in een hyperbolische curve, en dat PAR-niveaus worden uitgedrukt als percentages. Van Eyal et al. (2015).
In de Rode Zee en rond Hawaï, met zeer helder water, is de maximale diepte van zoöxanthellaatkoralen veel lager, namelijk 145-165 m (483-550 ft.) In vergelijking met gebieden met troebelere wateren zoals Curaçao (80 m of 267 ft.) ) en Oost-Florida (40 m of 133 ft, Kahng et al. 2010).

In tegenstelling tot wat vaak wordt gedacht, kan Acropora spp. groeien bij weinig licht. In de diepwateromgeving passen koralen en zoöxanthellen hun fysiologie en morfologie aan om het schaars beschikbare blauwe licht efficiënt op te vangen.
Het is bekend dat steenkoralen zich kunnen aanpassen aan lage lichtniveaus in aquaria, hoewel hun groeisnelheid en kleuring worden beïnvloed (Wijgerde en Laterveer 2013; Wijgerde en Tilstra 2014). In het veld zijn soortgelijke waarnemingen gedaan, met het recordhoudende zoöxanthellate steenkoraal Leptoseris hawaiiensis dat tot een diepte van minstens 165 meter (550 voet) is gevonden (Maragos en Jokiel 1986). Op zijn maximale recorddiepte ontvangt deze koraalsoort slechts 0,02% van de zonnestraling aan het oppervlak, of tussen 0 en 0,5 µmol fotonen m-2 s-1. Dit komt overeen met een slecht verlichte gang ‘s nachts. Om te overleven in deze donkere omgeving hebben diepwaterkoralen verschillende aanpassingen ondergaan. Ten eerste produceren hun endosymbiotische zoöxanthellen meer chlorofyl en andere fotopigmenten om de efficiëntie van lichtgebruik te verbeteren (Frade et al. 2008; Lesser et al. 2010). Bovendien veranderen de koralen zelf hun morfologie, met zijdelings afgeplatte takken, plaatachtige groeivormen en verder uit elkaar geplaatste korallieten, die hoogstwaarschijnlijk meer licht verzamelen (Lesser et al. 2010; Muir et al. 2015). Het koraalskelet fungeert als een efficiënte lichtcollector en verstrooit het licht op zo’n manier dat de zoöxanthellen het effectief kunnen gebruiken (Enriquez et al. 2005).
In diepere wateren produceren koralen vaak meer fotopigmenten zoals chlorofylen om licht efficiënter te vangen. Let op het bruine plaatkoraal Pachyseris speciosa in het midden en de bruinachtig gepigmenteerde toppen van de Seriatopora hystrix-kolonie rechtsonder, beide indicatief voor een verhoogde chlorofyl-a-synthese.
Naast bestraling verandert het lichtspectrum met de waterdiepte. Hoe dieper we de waterkolom induiken, hoe meer natuurlijk zonlicht wordt aangepast door de selectieve filterende eigenschappen van zeewater. Rood licht, met de grootste golflengte en de laagste energiefotonen binnen het zichtbare spectrum, wordt snel verzwakt door zeewater. Het dringt niet meer dan ongeveer 10 meter (33 voet) door in tropisch zeewater, hoewel deze waarde varieert met de locatie en tijd (Mass et al. 2010). Op 40 meter diepte en lager dringt licht met golflengten van 600 nm en verder (d.w.z. oranje, rood en infrarood) niet door (Rivero-Calle et al.2008; Mass et al.2010). Zo werkt zeewater als een chromatisch filter, waarbij blauw en groen dominant worden in diepere wateren.
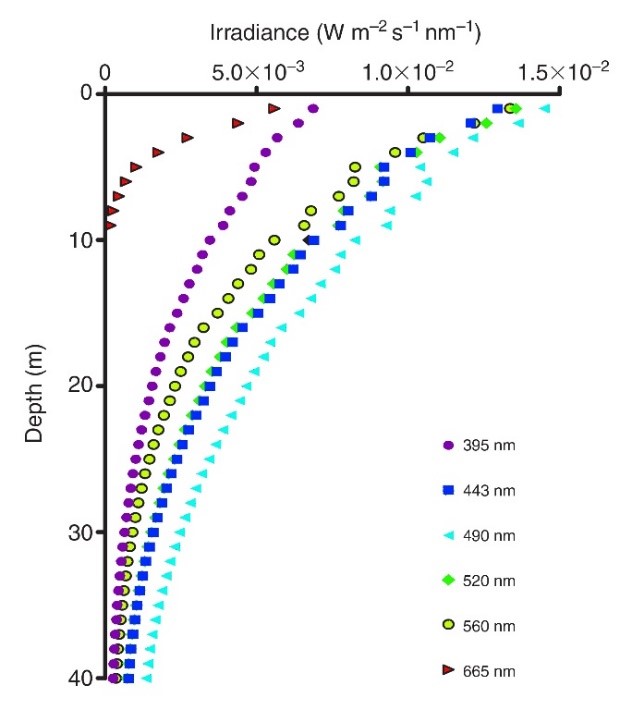
Bestralingsprofielen (395-665 nm) gemeten in koraalrifwateren in Eilat, Israël. Rode en oranje (niet weergegeven) golflengten dringen niet diep door in het zeewater, wat resulteert in blauwgroen water van meer dan 10 meter diepte. Aangepast van Mass et al. (2010).
Aanpassing aan een overwegend blauwe omgeving kan verklaren waarom blauw licht alleen voldoende lijkt om gezonde koralen en zoöxanthellen in aquaria te behouden, de laatste zowel in hospite (die van nature in het koraal groeit) als ex hospite (levend buiten het koraal in kweekflessen, Kinzie et al. 1984, 1987; Wang et al. 2008; Wijgerde et al. 2014). Koralen uit diepere wateren gebruiken blauw licht ook effectiever voor fotosynthese in vergelijking met ondiepe koralen, wat duidt op een fenomeen dat bekend staat als chromatische aanpassing, d.w.z. aanpassing aan een specifiek lichtspectrum of kleur (Mass et al.2010). Deze aanpassing kan plaatsvinden via verhogingen van de synthese van verschillende fotopigmenten door zoöxanthellen, zoals chlorofyl a en het carotenoïde peridinine.
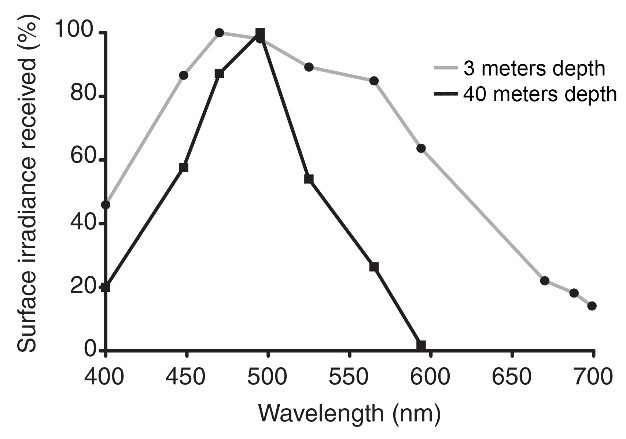
Op 3 meter diepte dringen alle kleuren door. Op 40 meter diepte is licht met golflengten boven 600 nm (oranje, rood en infrarood) echter volledig afwezig. Interessant is dat cyaanlicht (490 nm) zijn kracht blijft op 40 meter diepte, hoewel dit afhankelijk is van de locatie en de tijd van het jaar. Aangepast van Mass et al. (2010).

Een paar koraalvlinders (Chaetodon lunula) zweeft over een tonvormige spons.

Door digitale loggers in diepe wateren te plaatsen met behulp van duikboten, kunnen wetenschappers fluctuaties in temperatuur, lichtintensiteit en andere abiotische factoren op mesofotische riffen registreren. Let op de zweepkoralen op de achtergrond. Locatie: Curaçao.
Temperatuur
De watertemperatuur rond koraalriffen neemt af met de diepte, hoewel deze relatie sterk wordt beïnvloed door de locatie en de tijd van het jaar. Rond de Florida Keys, bijvoorbeeld, vertoont de watertemperatuur een scherpe daling van ongeveer 5 ° C (9 ° F) tussen 25 en 50 meter (83 tot 167 voet). Verderop daalt het langzaam naar lagere temperaturen in dieper water, met een kille 12 ° C (54 ° F) op 120 meter (400 voet) diepte. Hoewel andere locaties en seizoenen hogere temperaturen laten zien in diepere wateren (Rooney et al. 2010), laten metingen zien dat mesofotische riffen kunnen worden blootgesteld aan een breed temperatuurbereik van ongeveer 12 tot 29 ° C (54 tot 84 ° F). Dit zou (gedeeltelijk) het voorkomen van verschillende soorten over een dieptegradiënt kunnen verklaren, aangezien niet alle koralen dergelijke extreme temperaturen kunnen verdragen. Het toont ook aan dat veel tropische steenkoralen zich kunnen aanpassen aan zowel lage temperaturen als weinig licht. Interessant is dat mesofotische riffen tijdens zomerperiodes minder worden beïnvloed door hoge temperaturen, waardoor ze zelfs kunnen gedijen tijdens massale blekingsepisodes op ondiepe riffen (Lesser al. 2009).

In de sombere diepten van Curaçao inspecteert Dr. Norbert Englebert een grote spons.
Waterstroom
Naast instraling en temperatuur neemt de waterstroom ook af met de diepte, aangezien wind- en getijdekrachten alleen inwerken op de bovenste oceanische laag. Een uitzondering hierop vormen orkanen, die oceanische wateren kunnen mengen tot minstens 200 meter diep (Lesser et al. 2009). Mesfotische riffen leven dus over het algemeen in een omgeving met weinig energie, met verminderde waterstroom rond koralen. Een lage waterstroom, in combinatie met lage lichtniveaus, resulteert in morfologische aanpassingen door koralen zoals hierboven beschreven. Naast het vormen van plaatachtige morfologieën, worden koralen fijner vertakt, met dunne en fragiele kolonies (Kahng et al. 2010; Done 2011). Dit kan te wijten zijn aan het ontbreken van een behoefte aan een robuust skelet bij afwezigheid van sterke golfwerking.

Steenkoralen zoals Acropora spp. veranderen hun morfologie met diepte. (A) Diepzeespecialisten A. pichoni verzameld op 40 m en (B) A. tenella verzameld op 52 m, (C) diepte generalist A. echinata verzameld op 40 m, (D) typische dikke morfologie voor de ondiepe intergetijdende soorten A. humilis, verzameld op 1 m, (E) diepte generalist A. granulosa vanaf 46 m diepte en (F) vanaf 7 m diepte. Hertshoornkoralen verzameld op een diepte van minder dan 40 m hebben over het algemeen lateraal afgeplatte takken, een lichter en kwetsbaarder skelet en een grotere afstand tussen corallieten en takken. Schaalbalken: 1 cm. Van Muir et al. (2015).
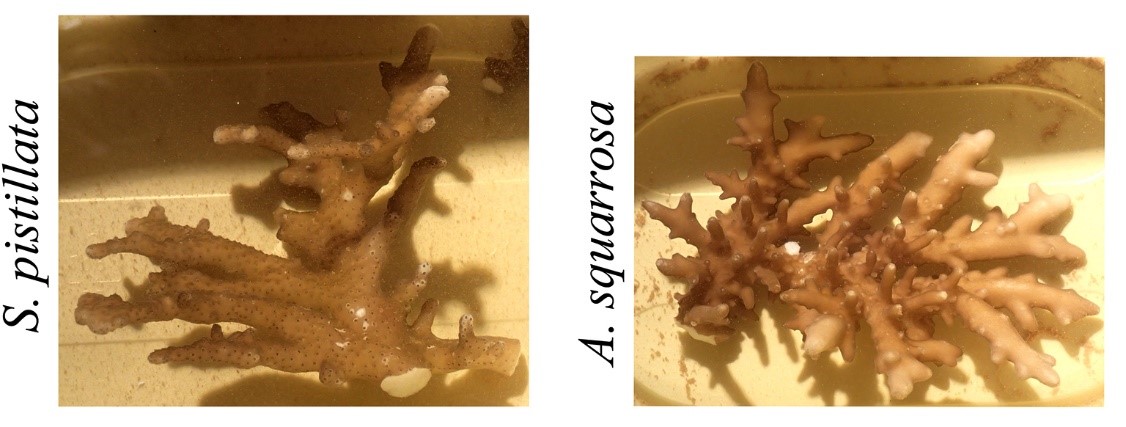
Wanneer koralen zich aanpassen aan zeer lage lichtniveaus, kan hun morfologie zo dramatisch worden veranderd dat het identificeren van soorten een uitdaging kan zijn. Deze foto’s tonen daglichtverschijningen van Stylophora pistillata en Acropora squarrosa verzameld op een diepte van ~ 60 m (200 ft). Let op de dunne en afgeplatte takken. Aangepast van Eyal et al. (2015).
Voedingsstoffen
Ondiep rifwater staat erom bekend uitzonderlijk arm te zijn aan opgeloste voedingsstoffen, met name nitraat en fosfaat (Lesser et al. 2009). Daarentegen vertonen diepere wateren verhoogde nutriëntenniveaus.
De nitraatconcentratie rond de Florida Keys is bijvoorbeeld ongeveer 0,1 µmol L-1 (0,006 mg L-1 of ppm) aan het oppervlak en neemt geleidelijk toe tot ongeveer 24 µmol L-1 (1,49 mg L-1) onder 200 meter diepte (Lesser et al.2009). Deze toename van nitraat met de diepte wordt veroorzaakt door opwelling, een fenomeen waarbij nutriënten vanuit de diepte naar het oppervlaktewater worden getransporteerd. Opwelling stimuleert de groei van fytoplankton (algen en cyanobacteriën), wat wordt gemeten door chlorofyl a, een fotosynthetisch pigment. In de Keys resulteert het opwellen van voedingsstoffen in een chlorofylpiek van ongeveer 60 meter (200 voet).
Hoewel mesofotische koralen kunnen worden aangepast aan eutrofisch (voedselrijk) water, hebben experimenten door wetenschappers en aquarianen in de loop der jaren aangetoond dat de meeste, zo niet alle koralen verhoogde nitraatgehaltes verdragen. Hoewel de groei van koraal in het algemeen wordt verminderd bij hogere nitraat- en fosfaatgehaltes (Marubini en Davies 1996; Ferrier-Pagès et al. 2000; Tanaka et al. 2007; Hylkema et al. 2014), kunnen mesofotische koralen uiteindelijk door beschikbaarheid en sedimentatie van licht in groei worden beperkt.
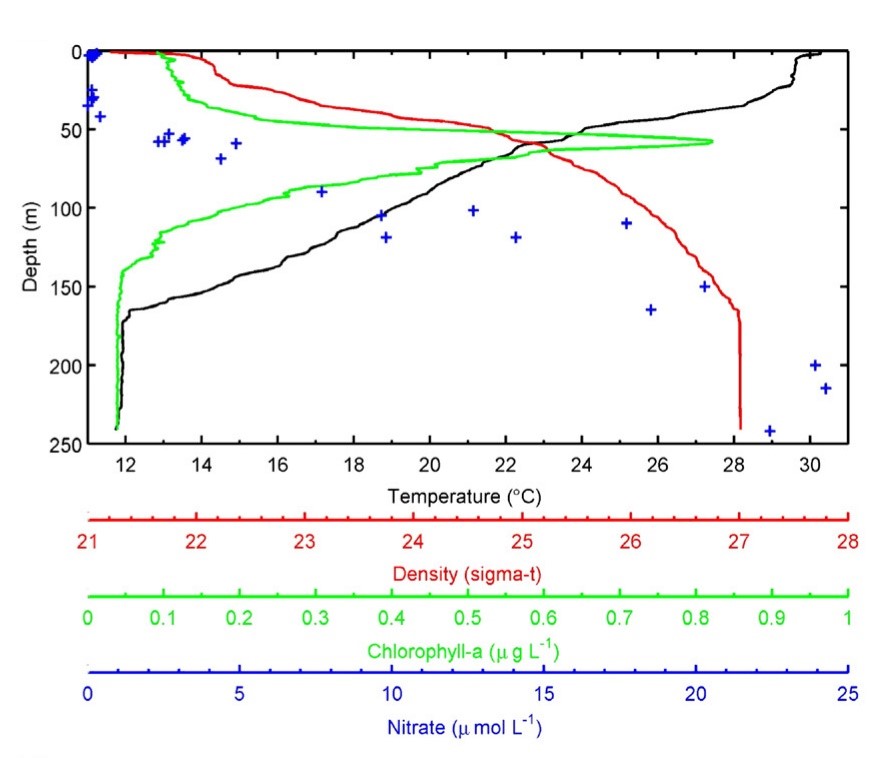
Profielen van temperatuur, waterdichtheid, chlorofyldichtheid en nitraatconcentratie met toenemende diepte verzameld bij Florida Keys. Merk op dat deze relaties sterk worden beïnvloed door locatie en seizoen, en worden als voorbeeld getoond. Gebaseerd op Lesser et al. (2009) en verwijzingen daarin.
Fluorescentie van koraal en vissen in de mesofotische zone
De fluorescerende aard van veel koralen is bekend bij zowel wetenschappers als aquarianen, waarbij één golflengte (kleur) licht wordt geabsorbeerd door fluorescerende eiwitten en wordt uitgezonden als een andere, maar altijd langere golflengte met een lagere energie en frequentie. Als ze bijvoorbeeld worden opgewekt door blauw licht, fluoresceren veel koralen dit licht als licht met een langere golflengte, in de vorm van groene, gele, oranje en rode kleuren. Hoewel fluorescentie zeer aantrekkelijk is, blijft de functie ervan onduidelijk. In de literatuur zijn verschillende biologische verklaringen voor koraalfluorescentie voorgesteld. Fluorescentie kan koralen beschermen tegen schadelijk licht in ondiep water door energetisch blauw licht te fluoresceren als minder schadelijk groen licht en door zuurstofradicalen op te vangen als bijproducten van fotosynthese (Bou-Abdallah et al. 2006; D’Angelo et al. 2008, 2012 ; Gittins et al.2015; Salih et al.2000). Bovendien kan het koralen helpen om in dieper water te overleven door het oogsten van licht te verbeteren (Eyal et al.2015).
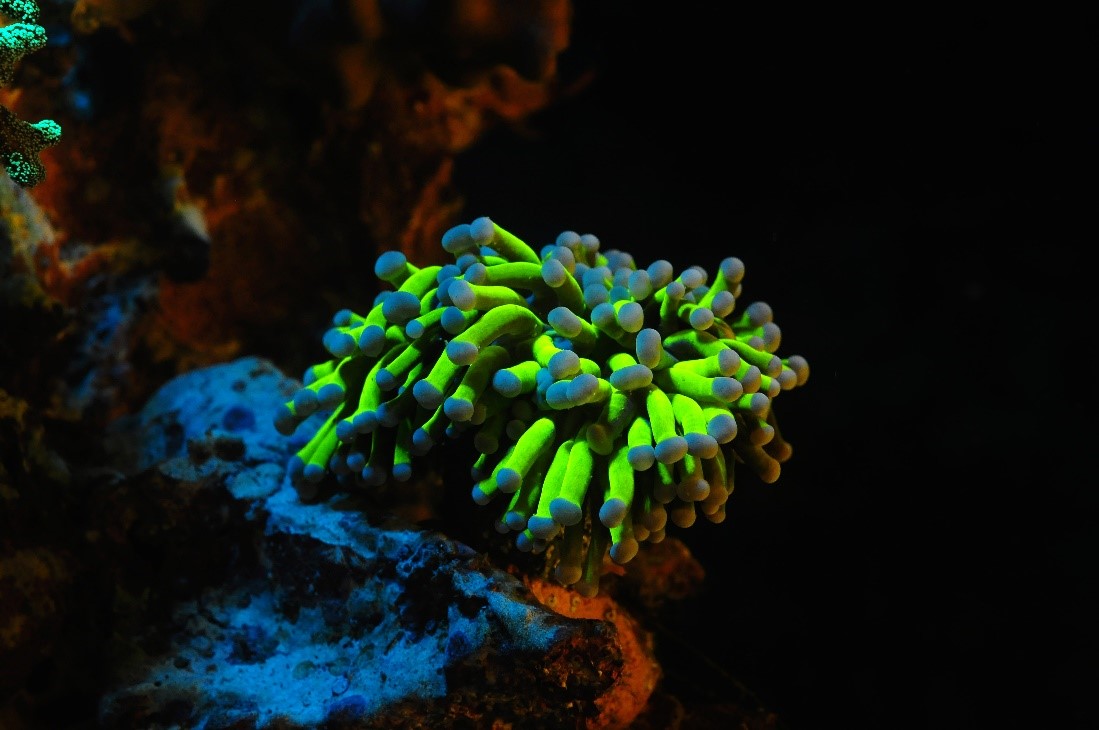
Gele fluorescentie is zeldzaam bij ondiepe koraalsoorten, en wordt vaker aangetroffen in diep water exemplaren. Omgekeerd is cyaanfluorescentie ongebruikelijk in diepwaterkoralen en wordt het vaak aangetroffen bij ondiepe soorten. Het is onduidelijk waarom. Deze Euphyllia sp. werd gefotografeerd met excitatielicht van 400-450 nm en een Cokin geel langdoorlaatfilter. Afbeelding door Tim Wijgerde.
Interessant is dat ondiepe rifkoralen verschillende fluorescentiepatronen vertonen in vergelijking met die uit de mesofotische zone. Oranjerode fluorescentie is bijvoorbeeld bijna afwezig in koralen van ondiepe riffen in Eilat, en komt veel vaker voor bij exemplaren die onder de 40 meter worden gevonden (Eyal et al. 2015). Bovendien behouden diepwaterkoralen die groene fluorescentie vertonen hun fluorescentie-intensiteit ongeacht de lichtniveaus, in tegenstelling tot ondiepwaterkoralen die sterkere fluorescentie vertonen bij hogere lichtintensiteit (Leutenegger et al.2007; Oswald et al.2007; Eyal et al. 2015). Dit suggereert dat fluorescentie, in het bijzonder als oranjerood licht, een nog onbekende biologische functie heeft in diepere wateren. Mogelijke rollen voor fluorescerende eiwitten in diepe en/of weinig licht omgevingen zijn het verbeteren van de fotosynthese (Schlichter et al. 1986; Salih et al. 2000), en visuele aanwijzingen voor symbiotische vissen (Matz et al. 2006) en camouflage tegen bepaalde vissen ( Matz et al. 2006). Verbetering van fotosynthese door fluorescerende eiwitten wordt ondersteund door hun lokalisatie in het endoderm van het koraal (onderste weefsellaag), onder zoöxanthellen bij weinig licht, waardoor deze eiwitten het licht kunnen reflecteren en fluoresceren naar de zoöxanthellen (Salih et al. 2000). Omgekeerd bevinden fluorescerende eiwitten zich onder omstandigheden met veel licht in het ectoderm (bovenste weefsellaag), boven de zoöxanthellen, waardoor deze pigmenten de hoeveelheid licht die de zoöxanthellen bereikt, kunnen verminderen (Salih et al. 2000). Cyaanfluorescentie is tenslotte ongebruikelijk in diepwaterkoralen, wat logisch lijkt omdat de fotoprotectieve functie niet nuttig is in een omgeving met weinig licht. Ondiepe koralen kunnen dit pigment gebruiken om overtollig zichtbaar licht te reflecteren en UV-licht te fluoresceren als blauw licht, waardoor hun weefsels en symbiotische zoöxanthellen worden beschermd tegen lichtschade (D’Angelo et al. 2008; Salih et al. 2000).
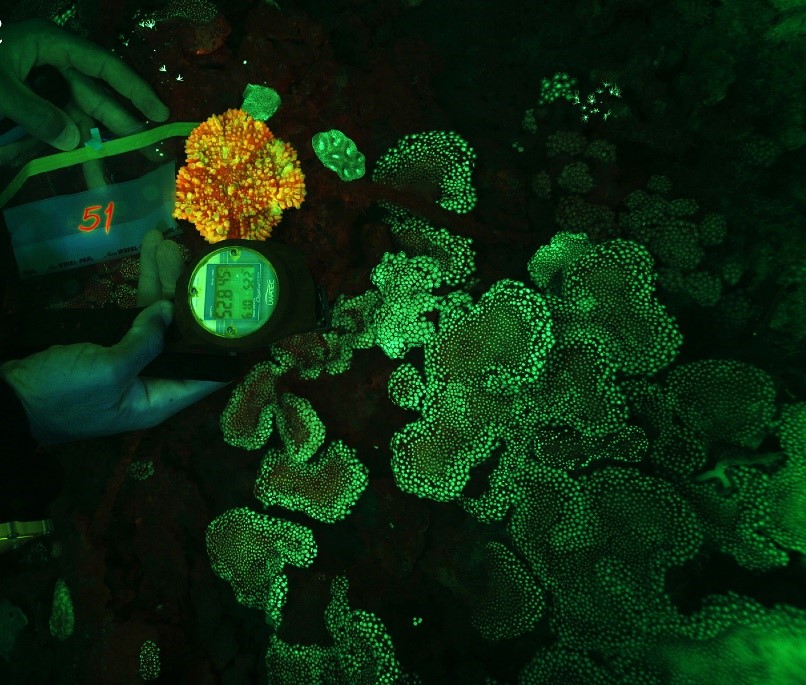
Oranjerode fluorescentie van Echinophyllia aspera en groene fluorescentie van Alveopora ocellata op 52,8 m (176 voet) in Eilat, Israël. Afbeelding aangepast van Eyal et al. (2015).
Hoewel koralen tot de meest fluorescerende dieren op het rif behoren, vertonen veel andere organismen deze eigenschap. Deze omvatten veel vissen, met name kleine en cryptische soorten. Onlangs ontdekten biologen dat leden van de GOBIIDAE, SYNGNATHIDAE en TRIPTERYGIIDAE opvallende rode fluorescerende ogen hebben (Meadows et al. 2014). Interessant is dat de intensiteit van oogfluorescentie toeneemt met de diepte, wat suggereert dat het enig voordeel oplevert in somber, blauw water. Door de rode gloed rond hun ogen kunnen deze vissen prooien gemakkelijker detecteren, potentiële partners signaleren en/of rivalen in mesofotische habitats waarschuwen.
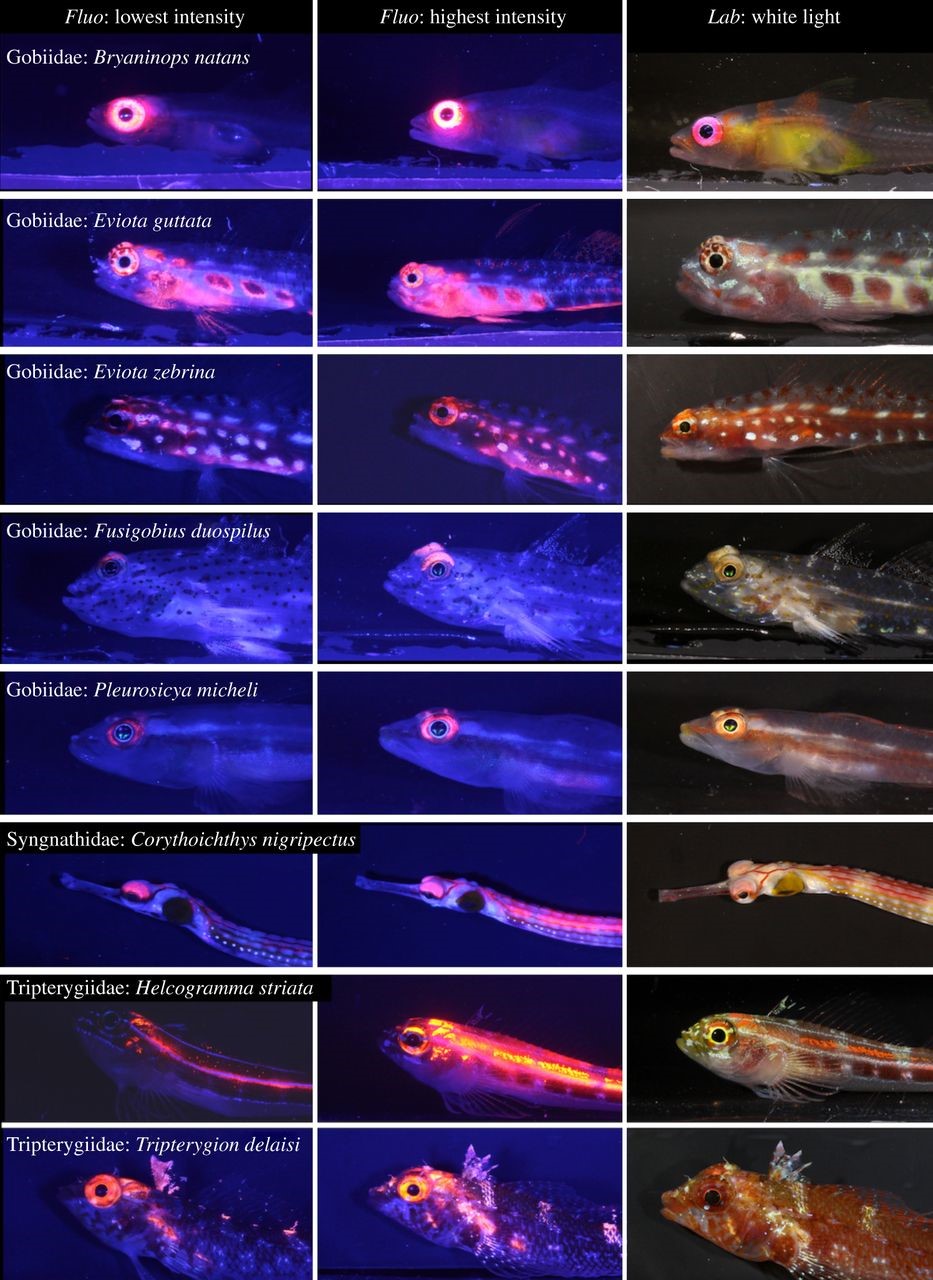
Veel kleine rifvissen vertonen rode fluorescentie rond hun ogen, met verhoogde intensiteit op lagere diepten. In diepe wateren kan de rode gloed rond hun ogen ervoor zorgen dat vissen prooien en signaalgenoten beter kunnen detecteren. De linker en middelste kolommen tonen foto’s die in het laboratorium onder blauw licht zijn genomen, met minimale en maximale fluorescentiehelderheid bij bemonsterde individuen. De rechterkolom toont vissen onder standaard witlichtomstandigheden. Alle vissen komen voor op koraalriffen, met uitzondering van de mediterrane soort Tripterygion delaisi. Van Meadows et al. (2014).
Plankton en heterotrofe voeding in de mesofotische zone
Het is bekend dat koralen zich voeden met plankton, detritisch materiaal en opgelost organisch materiaal, een fenomeen dat bekend staat als heterotrofie (besproken door Houlbrèque en Ferrier-Pagès 2009; Ferrier-Pagès et al. 2011; Wijgerde 2013). Heterotrofe voeding is een belangrijke strategie om voedingsstoffen te verkrijgen ter aanvulling van fotosynthese (autotrofie), en dit zou vooral nuttig zijn om te overleven in een habitat met weinig licht waar fotosynthese minder effectief is. Er is echter geen sluitend bewijs dat steenkoralen met symbiotische zoöxanthellen hun voedingssnelheid verhogen wanneer ze groeien in de mesofotische zone.
Door stabiele isotopen van koolstof en stikstof te analyseren, kunnen mariene biologen een specifieke chemische signatuur detecteren die onthult hoe koralen hun voedsel verkrijgen. Voor Montastraea cavernosa heeft isotopenanalyse van zijn weefsel, skelet en zoöxanthellen aangetoond dat de fotosynthese significant afneemt in diepere wateren. Hoewel Lesser et al. (2010) suggereerden dat op een diepte tussen 45 en 61 m het voeden met plankton en andere deeltjes de dominante voedingsmodus wordt, een meer diepgaande isotopenanalyse onthulde dat de voedingssnelheden vergelijkbaar blijven terwijl de fotosynthese afneemt (Crandall et al. 2016). Daarentegen gebruiken koralen van het genus Agaricia voeding als een primaire manier om voedingsstoffen te verwerven, ongeacht de diepte (Crandall et al.2016).
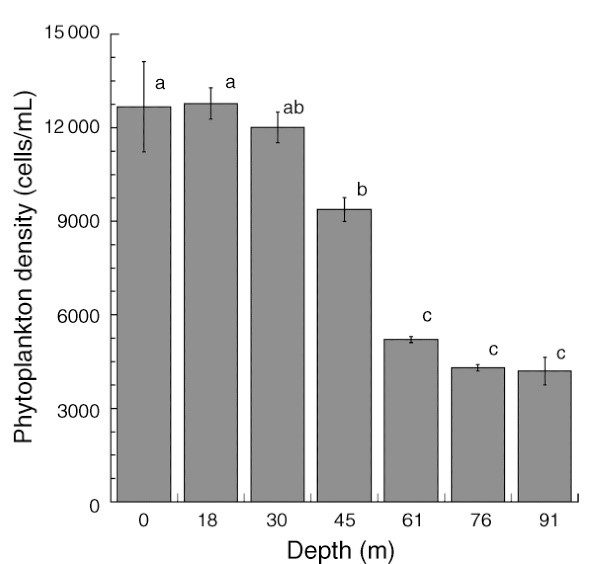
Totaal fytoplankton (algen en cyanobacteriën) beschikbaar vanaf het oppervlak tot 91 m diepte bij Bock Wall, Lee Stocking Island, Bahama’s. Tussen 30 en 61 meter neemt de dichtheid van fytoplankton aanzienlijk af, met gevolgen voor de koraalvoeding. Aangepast van Lesser et al. (2010).

Een zeebioloog inspecteert een mesofotisch koraalrif in Sulawesi (Celebes), dat een ongelooflijke biodiversiteit vertoont.
Aangezien de dichtheid van fytoplankton afneemt met de diepte (Lesser et al. 2010), hoogstwaarschijnlijk als gevolg van een verminderde beschikbaarheid van licht, zou het voor koralen moeilijker zijn om zich met deze deeltjes te voeden. Evenzo zou de mogelijke reductie van zoöplankton, dat grotendeels afhankelijk is van fytoplankton, de voedingssnelheid verder kunnen verlagen, ook al is zoöplankton aanwezig op een diepte van minstens 150 meter of 500 voet (Lesser et al. 2010). Door de beschikbaarheid van heterotrofe bacteriën, protisten en afval (zeesneeuw) in diepere wateren kunnen koralen nog steeds hun voedingssnelheid verhogen. Bovendien kan het opwellen van voedingsstoffen, zoals gemeten in de Florida Keys, resulteren in planktonbloei in diepere wateren, wat een voedselbron zou kunnen zijn voor mesofotische koralen.
Soorten, adaptieve divergentie en soortvorming
Zoals hierboven vermeld, hebben mesofotische riffen verschillende soortensamenstellingen in vergelijking met hun ondiepe tegenhangers. Groeien in de sombere diepten vereist aanzienlijke morfologische en fysiologische aanpassingen, en het lijkt erop dat niet alle soorten dit kunnen. Interessant is dat koraalsoorten die over een breed dieptebereik voorkomen (diepte-generalisten) uiteindelijk kunnen evolueren tot meerdere soorten als gevolg van DNA-mutaties en daaropvolgende natuurlijke selectie. Hieronder leg ik uit hoe dit werkt.
De vorming van nieuwe soorten staat bekend als soortvorming, en het begint met willekeurige chemische veranderingen die bekend staan als mutaties in het DNA - dat alle erfelijke informatie in levende cellen bevat - die in elk individu voorkomen. Soms heeft een dergelijke mutatie een gunstig effect, omdat het een gen - een DNA-fragment dat codeert voor een eiwit met een specifieke functie in een organisme - zodanig verandert dat een individu zich beter aanpast aan zijn omgeving. Een genetische verandering die resulteert in een efficiënter gebruik van zonlicht kan bijvoorbeeld een koraal individu in staat stellen om in dieper water te overleven. Langere overleving zal op zijn beurt het koraal in staat stellen om meer nakomelingen (seksuele voortplanting) of klonen te produceren door fragmentatie (aseksuele voortplanting), wat resulteert in meer koralen met dit gunstige gen (genotype) en bijbehorende fysieke kenmerken (fenotype). Dit proces, waarbij beter aangepaste individuen hogere overlevings- en reproductiesnelheden vertonen, ook wel bekend als een hogere conditie, wordt natuurlijke selectie genoemd. De combinatie van willekeurige DNA-mutaties en daaropvolgende niet-willekeurige natuurlijke selectie op die mutaties resulteert in evolutie, wat een geleidelijke verandering is in de vorm en functie van organismen.

Anthiassen en juffers verstoppen zich in de diepte.
Uiteindelijk kan een reproductief geïsoleerde koraalpopulatie ontstaan, als planula (koraallarven) van ondiep groeiende koralen moeite kunnen hebben om te overleven in een diepe, weinig lichtomgeving, en vice versa. Als dit gebeurt, kan de ondiepe populatie op een gegeven moment niet langer kruisen met de diepe populatie, omdat ze genetisch en fysiek te verschillend zijn geworden. Tegen die tijd, wanneer nakomelingen van diepe en ondiepe populaties in hoofdzaak onvruchtbaar zouden blijken te zijn, zoals de muilezel, zouden deze koraalpopulaties verschillende soorten worden genoemd en zouden hun onvruchtbare nakomelingen als hybriden worden beschouwd. De lange genetische weg naar soortvorming, veroorzaakt door willekeurige DNA-mutaties en de daaropvolgende natuurlijke selectie van die mutaties die individuen beter aanpassen aan een habitat, wordt adaptieve divergentie genoemd. Er zijn inderdaad aanwijzingen dat dit fenomeen voorkomt in koralen, zoals in het Indo-Pacifische koraal Seriatopora hystrix (Bongaerts et al. 2011a).
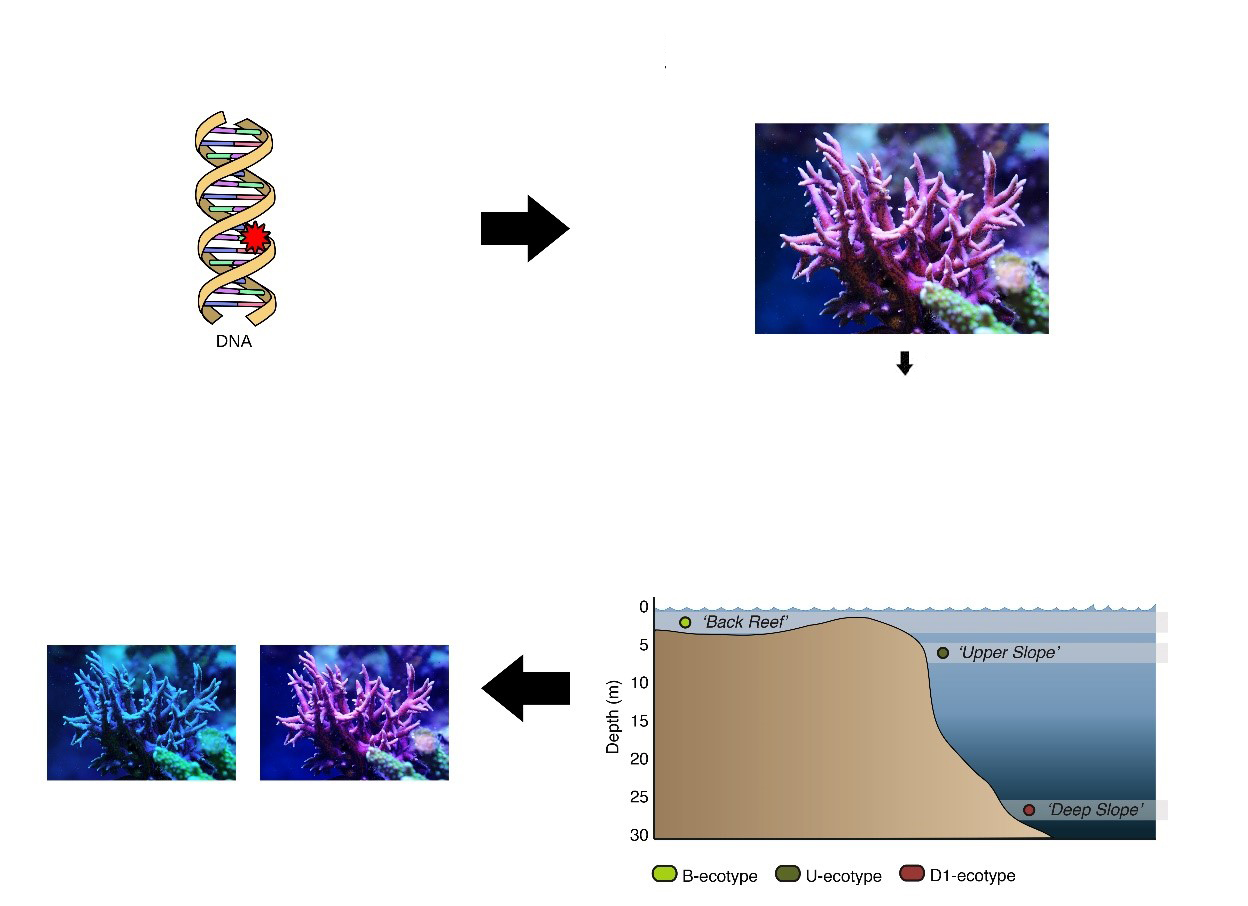
Schematisch overzicht van hoe een koraal zoals Seriatopora hystrix langzaam zou kunnen evolueren naar ondiepe en diepe soorten. Afbeeldingen door Tim Wijgerde, Forluvoft / Wikimedia Commons en Bongaerts et al. (2011a).
Voor Seriatopora hystrix is onlangs aangetoond dat er verschillende zogenaamde ecotypen bestaan, populaties van verschillende diepten met significante genetische en fysieke variaties daartussen (Bongaerts et al. 2011a). Deze verschillen werden ook gevonden tussen de symbiotische zoöxanthellen van de verschillende ecotypes, wat duidt op natuurlijke selectie op een zogenaamd holobiont-niveau, d.w.z. het koraal en de bijbehorende symbiotische organismen. Deze ecotypen, die hoogstwaarschijnlijk het gevolg zijn van adaptieve divergentie, kunnen bijna verschillende soorten worden. Dit werd aangetoond door verschillende overlevingspercentages nadat alle drie de ecotypes waren getransplanteerd over verschillende dieptehabitats. Het meest opvallende was dat het diepe ecotype een hogere overleving vertoonde op 25 m (83 ft) in vergelijking met ondiepe ecotypes wanneer deze tot dezelfde diepte werden getransplanteerd. Omgekeerd kon het diepe ecotype niet overleven tussen 0 en 5 m (0-17 ft), in tegenstelling tot de ondiepe ecotypes. Aangezien het diepe ecotype geen hogere fotosynthetische efficiëntie vertoonde dan de ondiepe ecotypen, zou zijn hogere overleving in dieper water te wijten kunnen zijn aan een verbeterd vermogen om zich te voeden met plankton (Bongaerts et al. 2011a). Dit moet echter nog worden bepaald. Hoewel diepzeekolonies tijdens dit experiment niet in ondiepe wateren overleefden, zijn er aanwijzingen dat op sommige riffen S. hystrix-larven hun weg terug naar de oppervlakte vinden en kunnen helpen bij het herbevolken van beschadigde riffen (van Oppen et al. 2011).
Morfologische en fysiologische aanpassing van een koraalpopulatie aan een specifieke omgeving, veroorzaakt door natuurlijke selectie die de zwakkere (minder aangepaste) individuen uitwist, heeft opmerkelijke effecten. Het meest opvallend voor aquariumliefhebbers is misschien wel de maximale diepte waarop zoöxanthellaatkoralen worden gevonden. Acropora spp., In hobbyistenkringen beschouwd als “licht veeleisend”, blijken een diepte van ten minste 73 m (243 ft) te hebben. Hier zijn de maximale PAR-niveaus ongeveer 30 µmol fotonen m-2 s-1, of 1-2% van de maximale zonnestraling (Lesser et al. 2010). Dit zou een laag lichtniveau in een aquarium betekenen. Hoewel hun groeisnelheid, morfologie en kleuring allemaal worden beïnvloed, is het interessant om te zien dat dergelijke koralen in diepe wateren groeien.
Van alle diep-levende zoöxanthellaat-steenkoralen neemt Leptoseris hawaiiensis de leiding en groeit tot 165 meter diep, waar slechts 0,02% van het zonlicht doordringt! Andere diepe soorten zijn Agaricia grahamae op 119 m (397 ft) in het Caribisch gebied en Leptoseris fragilis op 145 m (483 ft) in de Rode Zee (Reed 1985; Maragos en Jokiel 1986; Fricke et al.1987).
Hoewel Leptoseris hawaiiensis de recordhouder is voor zoöxanthellaatkoralen, worden andere fotosynthetische organismen aangetroffen in nog extremere habitats. Koraalalgen met korstmos zijn de diepst voorkomende macroalgen en zijn tot op een duizelingwekkende diepte van 268 meter (893 voet) aangetroffen, waar licht vrijwel afwezig is. Het is raadselachtig om daar beneden te ontdekken wat wij beschouwen als lichtafhankelijke organismen. Koraalalgen kunnen zelfs tot 17 maanden onder ijs overleven met minder dan 0,07% van de oppervlakte-instraling (Schwarz et al. 2005). Zeegrassen worden ook aangetroffen in diepe wateren, met een maximale diepte tot nu toe op 90 m (300 ft.).

Onderzoek naar mesofotische riffen is technisch uitdagend, omdat het onderzeeërs, op afstand bediende voertuigen (ROV’s) en speciale duikuitrusting vereist. Hier verplanten Dr. Pim Bongaerts en zijn collega’s diepwaterkoralen voor een wetenschappelijk experiment.
Symbiose met ZOÖXANTHELLAE en bacteriën
Zoals ik hierboven heb beschreven, vertonen koralen die in diepere wateren groeien, blijk van genetische en fysiologische aanpassing. Dit geldt niet alleen voor de koraalgastheer zelf, maar ook voor zijn symbiotische dinoflagellaten, de zoöxanthellen. Bongaerts et al. (2011a) ontdekten dat diepe exemplaren van Seriatopora hystrix genetisch unieke zoöxanthellen bevatten, die niet worden aangetroffen in hun ondiepe tegenhangers. Deze zoöxanthellen behoren tot een specifieke groep Symbiodinium, ook wel een clade genoemd. Hoewel alle S. hystrix gevonden in ondiepe en diepe wateren clade C ZOÖXANTHELLAE herbergden, werden subtiele verschillen in hun DNA gedetecteerd die suggereren dat er verschillende clade C-subgroepen bestaan, die mogelijk zijn aangepast aan specifieke dieptehabitats.
Onderstaande figuur illustreert het ecotypeconcept, waarbij specifieke koraal- en zoöxanthellengenotypes samen verschillende ecotypes vormen. Bewijs voor aanpassing door zoöxanthellen aan diepere wateren wordt ook geleverd door het werk van Lesser et al. (2010), die ontdekten dat onder de 61 m de genetische samenstelling van zoöxanthellen verandert. Volgens hen “is er een sterke selectie op zoöxanthellen die geschikt zijn om te overleven in de lichtbeperkte omgeving.”

Een ROV, klaar om een koraalmonster te nemen in de mesofotische zone.

Verschillende kleine steenkoralen groeien op een diepe plaats rond Bunaken, Sulawesi. Let op de paarse sponzen en de manteldieren precies uit het midden, mogelijk Didemnum molle.

Doornenkroonzeester (Acanthaster planci) wordt ook aangetroffen in de mesofotische zone.

Dr. Pim Bongaerts gebruikt een op afstand bestuurd voertuig (ROV) om diepwaterkoralen op het Great Barrier Reef te bestuderen.

Een school doktersvissen doorkruist een mesofotisch rif rond Sulawesi.

Gewoon weer een dag op kantoor: een zeebioloog fotografeert een rijke diepwaterrifgemeenschap. Let op de tonvormige spons net onder de handen van de duiker en het spiraalvormige zwarte koraal rechts van hem.
Hoewel zoöxanthellen scleractinische koralen tot op een diepte van 165 meter (550 voet) van voeding kunnen voorzien, is dit voor zwarte koralen nog onduidelijk (orde ANTIPATHARIA). Hoewel antipatharians lange tijd als azoöxanthellate werden beschouwd, is onlangs gevonden dat ze Symbiodinium bevatten (Wagner et al. 2011). Zelfs op een diepte van ongeveer 400 meter (1333 voet) kunnen deze koralen Symbiodinium herbergen, waarvan de meeste behoren tot clade C. Aangezien zo’n diepe, donkere habitat de fotosynthese ondoelmatig maakt, kunnen veel zwarte koralen worden geparasiteerd door ZOÖXANTHELLAE, hoewel dit nog onderzocht moet worden.
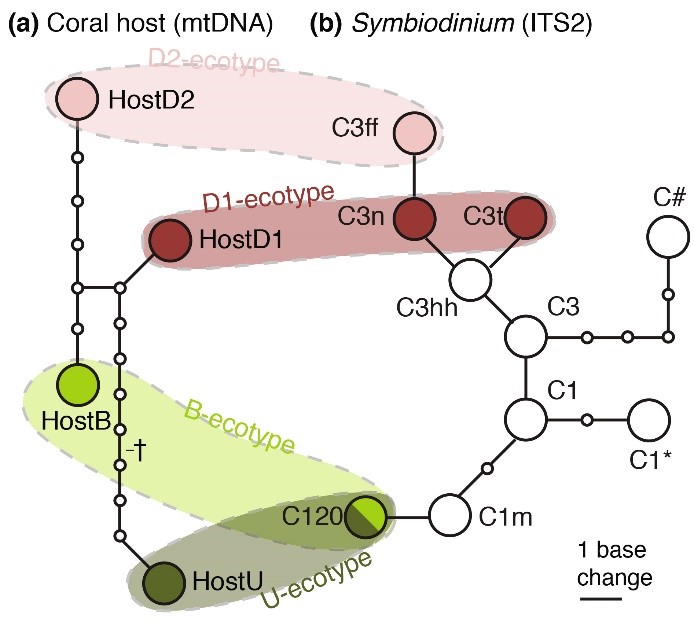
Op basis van DNA-markers (mitochondriaal DNA / mtDNA en nucleair DNA / ITS2) kunnen genetisch verschillende Seriatopora hystrix (a) en ZOÖXANTHELLAE (b) worden gedetecteerd, die genotypen worden genoemd. DNA-analyse van diepe en ondiepe S. hystrix-kolonies laten zien dat er specifieke associaties tussen koraalgenotypes en zoöxanthellengenotypes voorkomen. De vier gekleurde gebieden geven specifieke combinaties van koraal- en zoöxanthellengenotypes aan, die samen de verschillende ecotypen van Seriatopora hystrix vormen, namelijk de ecotypen Back Reef (B), Upper Slope (U) en Deep Slope (D1 / D2). Van Bongaerts et al. (2011a).
Naast zoöxanthellen heeft onderzoek aangetoond dat koralen gastheer zijn voor een diverse bacteriegemeenschap. Deze bacteriën leven in slijm op het oppervlak van het koraal, in koraalweefsel en zelfs in zoöxanthellen (Ainsworth et al.2015). Specifieke koraalsoorten associëren zich met bepaalde bacteriën, vergelijkbaar met zoöxanthellen (Olson en Kellogg 2010). Wat voor aquarianen misschien verrassend is, is dat bepaalde cyanobacteriën worden aangetroffen in koralen en hun zoöxanthellen, die stikstof binden door opgelost stikstofgas (N2) om te zetten in ammoniak, dat vervolgens wordt overgebracht naar de zoöxanthellen (Lesser et al. 2007). Deze gebruiken op hun beurt de ammoniak om aminozuren en eiwitten te produceren en zichzelf en hun koraalgastheer te voeden. Deze ingewikkelde samenwerking tussen het koraaldier en zijn symbiotische micro-organismen zorgt ervoor dat deze zogenaamde holobiont kan overleven in een omgeving met weinig voedingsstoffen. Hoewel bepaalde zoöxanthellen alleen op diepere koraalriffen voorkomen, is dit voor bacteriën nog niet duidelijk (Olson en Kellogg 2010). Het is mogelijk dat specifieke bacteriesoorten ervoor zorgen dat koralen blijven bestaan in omgevingen met zeer weinig licht, door hun gastheer te voorzien van specifieke verbindingen.

Zelfs een klein stukje mesofotisch rif zit boordevol biodiversiteit. Let op de kleurrijke kantkoralen, mogelijk Stylaster of Distichopora sp. Kantkoralen behoren tot de Hydrozoa-klasse en produceren een calciumcarbonaatskelet, waardoor ze lijken op steenkoralen (Anthozoa-klasse). Ze planten zich voort door miniatuurkwallen in het water vrij te laten.

Antipatharia, in de volksmond bekend als zwarte koralen, gedijen goed in de diepe wateren van Sulawesi. Hier overleven ze door plankton en afval te vangen. De rol van hun endosymbiotische zoöxanthellen blijft onduidelijk.

Het analyseren van slijm levert belangrijke aanwijzingen op over het leven van een koraal in de diepte, zoals de associaties met specifieke bacteriën. Hier bemonstert Dr. Pedro Frade een steenachtig koraal dat is verzameld vanaf 90 meter (300 voet).

Mesofotische riffen, de thuishaven van een fascinerend onderwaterleven, verdienen onze bescherming. Let op de Anthias die net uit het midden is vertrokken.
Afsluitende opmerkingen
Het is duidelijk dat mesofotische koraalriffen fascinerende, belangrijke en ook ondergewaardeerde ecosystemen zijn. Gelukkig wint de wetenschappelijke studie van mesofotische riffen aan grip, waardoor we een beter begrip krijgen van hoe deze ecosystemen functioneren. Openbare aquaria besteden er ook aandacht aan, zoals het Steinhart Aquarium van de California Academy of Sciences dat een mesofotisch rifaquarium voorbereidt, met koralen en vissen die speciaal voor deze nieuwe tentoonstelling zijn verzameld. Door onderzoek en onderwijs zullen we hopelijk in staat zijn om deze riffen te beschermen tegen menselijke verstoringen, zodat ze kunnen dienen als toevluchtsoord voor vissen, koralen en andere ongewervelde dieren.

Grote scholen blauwe trekkervissen (Odonus niger) vinden hun toevlucht in de diepte, tussen grote sponzen, zachte koralen en gorgonen. Let op de eenzame vissen net links en rechts uit het midden.
Verder lezen
Ga voor meer informatie over (mesofotische) koraalriffen naar de website van XL Catlin Seaview Survey, een uniek project dat tot doel heeft de riffen van de wereld vast te leggen in een panoramisch zicht van 360 graden. Met behulp van een innovatieve SVII panoramische camera met propeller en drie Canon 5D DSLR’s uitgerust met groothoeklenzen, worden 360 graden beelden van koraalriffen opgenomen. Hun website heeft een aantal spectaculaire afbeeldingen die u kunt downloaden. De gerelateerde XL Catlin Global Reef Record heeft ook veel panoramische afbeeldingen en video’s om van te genieten.

Dr. Pim Bongaerts bedient een camera met krachtige lichten op het Great Barrier Reef.
Erkenningen
Ik ben dr. Pim Bongaerts van het Global Change Institute, University of Queensland en zijn collega’s van de XL Catlin Seaview Survey enorm veel dank verschuldigd voor het leveren van buitengewone beelden. Ik wil ook Charles Delbeek, M.Sc., assistent-curator en Kelly Mendez, B.A., Senior Communications Manager van het Steinhart Aquarium bedanken voor het verstrekken van informatie over hun nieuwe mesofotische display.
Bronnen:
1. Ainsworth TD, Krause L, Bridge T, Torda G, Raina J-P, Zakrzewski M, Gates RD, Padilla-Gamiño JL, Spalding HL, Smith C, Woolsey ES, Bourne DG, Bongaerts P, Hoegh-Guldberg O, Leggat W (2015) The coral core microbiome identifies rare bacterial taxa as ubiquitous endosymbionts. The ISME Journal 9:2261-2274
2. Bak RPM, Nieuwland G, Meesters EH (2005) Coral reef crisis in deep and shallow reefs: 30 years of constancy and change in reefs of Curaçao and Bonaire. Coral Reefs 24:475-479
3. Bongaerts P, Ridgway T, Sampayo EM, Hoegh-Guldberg O (2010) Assessing the ‘deep reef refugia’ hypothesis: focus on Caribbean reefs. Coral Reefs 29(2):309-327
4. Bongaerts P, Riginos C, Hay KB, van Oppen MJH, Hoegh-Guldberg O, Dove S (2011a) Adaptive divergence in a scleractinian coral: physiological adaptation of Seriatopora hystrix to shallow and deep reef habitats. BMC Evolutionary Biology 11:303
5. Bongaerts P, Sampayo EM, Bridge TCL, Ridgway T, Vermeulen F, Englebert N, Webster JM, Hoegh-Guldberg O (2011b) Symbiodinium diversity in mesophotic coral communities on the Great Barrier Reef: a first assessment. Marine Ecology Progress Series 439:117-126
6. Bou-Abdallah F, Chasteen ND, Lesser MP (2006) Quenching of superoxide radicals by green fluorescent protein. Biochim Biophys Acta 1760:1690-1695
7. Bridge TCL, Fabricius KE, Bongaerts P, Wallace CC, Muir PR, Done TJ, Webster JM (2012) Diversity of Scleractinia and Octocorallia in the mesophotic zone of the Great Barrier Reef, Australia. Coral Reefs 31:179-189
8. Crandall JB, Teece MA, Estes BA, Manfrino C, Ciesla JH (2016) Nutrient acquisition strategies in mesophotic hard corals using compound specific stable isotope analysis of sterols. Journal of Experimental Marine Biology and Ecology 474:133-141
9. D’Angelo C, Denzel A, Vogt A, Matz MV, Oswald F, Salih A, et al. (2008) Blue light regulation of host pigment in reef-building corals. Marine Ecology Progress Series 364:97-106
10. D’Angelo C, Smith EG, Oswald F, Burt J, Tchernov D, Wiedenmann J (2012) Locally accelerated growth is part of the innate immune response and repair mechanisms in reef-building corals as detected by green fluorescent protein (GFP)-like pigments. Coral Reefs 31:1045-1056
11. De Goeij JM, van Oevelen D, Vermeij MJA, Osinga R, Middelburg JJ, de Goeij AFPM, Admiraal W (2013) Surviving in a Marine Desert: The Sponge Loop Retains Resources Within Coral Reefs. Science 342:108-110
12. Done T (2011) Corals: environmental controls on growth. In: Hopley D, editor. Encylopedia of modern coral reefs. Springer, Dordrecht. pp 281-293
13. Enriquez S, Mendez ER, Iglesias-Prieto R (2005) Multiple scattering on coral skeletons enhances light absorption by symbiotic algae. Limnology and Oceanography 50(4):1025-1032
14. Eyal G, Wiedenmann J, Grinblat M, D’Angelo C, Kramarsky-Winter E, Treibitz T, et al. (2015) Spectral Diversity and Regulation of Coral Fluorescence in a Mesophotic Reef Habitat in the Red Sea. PLoS ONE 10(6): e0128697. doi:10.1371/journal.pone.0128697
15. Ferrier-Pagès C, Gattuso JP, Dallot S, Jaubert J (2000) Effect of nutrient enrichment on growth and photosynthesis of the zooxanthellate coral Stylophora pistillata. Coral Reefs 19:103-113
16. Ferrier-Pagès C, Hoogenboom M, Houlbrèque F (2011) The role of plankton in coral trophodynamics, 215-229. In: Dubinsky Z, Stambler N (Eds), Coral reefs: an ecosystem in transition. Springer, Dordrecht, The Netherlands
17. Frade PR, Bongaerts B, Winkelhagen AJS, Tonk L, Bak RPM (2008) In situ photobiology of corals over large depth ranges: a multivariant analysis on the roles of environment, host, and algal symbiont. Limnology and Oceanography 53:2711-2723
18. Fricke HW, Vareschi E, Schlichter D (1987) Photoecology of the coral Leptoseris fragilis in the Red Sea twilight zone (an experimental study by submersible). Oecologia 73:371-381
19. Gattuso J-P, Gentilli B, Duarte CM, Kleypas JA, Middelburg JJ, Antoine D (2006) Light availability in the coastal ocean: impact on the distribution of benthic photosynthetic organisms and their contribution to primary production. Biogeoscience 3:489-513
20. Gittins JR, D’Angelo C, Oswald F, Edwards RJ, Wiedenmann J (2015) Fluorescent protein-mediated colour polymorphism in reef corals: multicopy genes extend the adaptation/acclimatization potential to variable light environments. Molecular Ecology 24: 453-465
21. Houlbrèque F, Ferrier-Pagès C (2009) Heterotrophy in tropical scleractinian corals. Biol Rev Camb Philos 84:1-17
22. Hylkema A, Wijgerde T, Osinga R (2015) Decreased growth of Stylophora pistillata with nutrient-driven elevated zooxanthellae density is largely explained by DIC limitation. Advanced Aquarist 14(5)
23. Kahng SE, Garcia-Sais JR, Spalding HL, Brokovich E, Wagner D, Weil E, Hinderstein L, Toonen RJ (2010) Community ecology of mesophotic coral reef ecosystems. Coral Reefs 29:255-275
24. Kahng SE, Maragos JE (2006) The deepest, zooxanthellate scleractinian corals in the world? Coral Reefs 25:254
25. Kinzie III RA, Hunter T (1987) Effect of light quality on photosynthesis of the reef coral Montipora verrucosa. Mar Biol 94:95-109
26. Kinzie III RA, Jokiel PL, York R (1984) Effects of light of altered spectral composition on coral zooxanthellae associations and on zooxanthellae in vitro. Mar Biol 78:239-248
27. Kühlmann DHH (1983) Composition and ecology of deep-water coral associations. Helgoländer Meeresuntersuchungen 36:183-204
28. Lesser MP, Falcón LI, Rodríguez-Román A, Enríquez S, Hoegh-Guldberg O, Iglesias-Prieto R (2007) Nitrogen fixation by symbiotic cyanobacteria provides a source of nitrogen for the scleractinian coral Montastraea cavernosa. Marine Ecology Progress Series 346:143-152
29. Lesser MP, Slattery M, Leichter JJ (2009) Ecology of mesophotic coral reefs. Journal of Experimental Marine Biology and Ecology 375:1-8
30. Lesser MP, Slattery M, Stat M, Ojimi M, Gates RD, Grottoli A (2010) Photoacclimatization by the coral Montastraea cavernosa in the mesophotic zone: light, food, and genetics. Ecology 91(4):990-1003
31. Leutenegger A, D’Angelo C, Matz MV, Denzel A, Oswald F, Salih A, et al. (2007) It’s cheap to be colorful. Anthozoans show a slow turnover of GFP-like proteins. FEBS Journal 274:2496-2505
32. Maragos JE, Jokiel PL (1986) Reef corals of Johnston Atoll-one of the World’s most isolated reefs. Coral Reefs 4:141-150
33. Marubini F, Davies PS (1996) Nitrate increases zooxanthellae population density and reduces skeletogenesis in corals. Mar Biol 127:319-328
34. Mass T, Kline DI, Roopin M, Veal CJ, Cohen S, et al. (2010) The spectral quality of light is a key driver of photosynthesis and photoadaptation in Stylophora pistillata colonies from different depths in the Red Sea. J Exp Biol 213:4084-4091
35. Matz MV, Marshall NJ, Vorobyev M (2006) Are Corals Colorful? Photochemistry and Photobiology 82:345-350
36. Meadows MG, Anthes N, Dangelmayer S, Alwany MA, Gerlach T, Schulte G, et al. (2014) Red fluorescence increases with depth in reef fishes, supporting a visual function, not UV protection. Proceedings of the Royal Society of London B: Biological Sciences 281: 20141211. http://dx.doi.org/10.1098/rspb.2014.1211
37. Muir P, Wallace C, Bridge TCL, Bongaerts P (2015) Diverse Staghorn Coral Fauna on the Mesophotic Reefs of North-East Australia. PLoS ONE 10(2): e0117933. doi:10.1371/journal.pone.0117933
38. Olson JB, Kellogg CA (2010) Microbial ecology of corals, sponges, and algae in mesophotic coral environments. FEMS Microbiology Ecology 73:17-30
39. Oswald F, Schmitt F, Leutenegger A, Ivanchenko S, D’Angelo C, Salih A, et al. (2007) Contributions of host and symbiont pigments to the coloration of reef corals. FEBS Journal 274:1102-1109
40. Reed JK (1985) Deepest distribution of Atlantic hermatypic corals discovered in the Bahamas. In: Proceedings of the 5th international coral reef symposium 6:249-254
41. Rivero-Calle S, Armstrong RA, Soto-Santiago FJ (2008) Biological and physical characteristics of a mesophotic coral reef: Black Jack reef, Vieques, Puerto Rico. Proceedings of the 11th International Coral Reef Symposium, Ft. Lauderdale, Florida, 7-11 July, Session number 16
42. Rooney J, Donham E, Montgomery A, Spalding H, Parrish F, Boland R, Fenner D, Gove J, Vetter O (2010) Mesophotic coral ecosystems in the Hawaiian Archipelago. Coral Reefs 29:361-367
43. Salih A, Hoegh-Guldberg O and Cox G (1998) Photoprotection of symbiotic dinoflagellates by fluorescent pigments in reef corals. Proc Aust Coral Reef Society 75 Anniv Conference (ed) Greenwood & Hall. pp 217-230
44. Salih A, Larkum A, Cox G, Kuhl M, Hoegh-Guldberg O (2000) Fluorescent pigments in corals are photoprotective. Nature 408:850-853
45. Schlichter D, Fricke HW, Weber W (1986) Light harvesting by wavelength transformation in a symbiotic coral of the Red Sea twilight zone. Marine Biology 91:403-407
46. Schwarz A-M, Hawes I, Andrew N, Mercer S, Cummings V, Thrush SF (2005) Primary production potential of nongeniculate coralline algae at Cape Evans, Ross Sea, Antarctica. Marine Ecology Progress Series 294:131-140
47. Tanaka Y, Miyajima T, Koike I, Hayashibara T, Ogawa H (2007) Imbalanced coral growth between organic tissue and carbonate skeleton caused by nutrient enrichment. Limnol Oceanogr 52:1139-1146
48. van Oppen MJH, Bongaerts P, Underwood JN, Peplow LM, Cooper TF (2011) The role of deep reefs in shallow reef recovery: an assessment of vertical connectivity in a brooding coral from west and east Australia. Molecular Ecology doi: 10.1111/j.1365-294X.2011.05050.x
49. Wagner D, Pochon X, Irwin L, Toonen RJ, Gates RD (2011) Azooxanthellate? Most Hawaiian black corals contain Symbiodinium. Proceedings of the Royal Society B 278:1323-1328
50. Wang L-H, Liu Y-H, Ju Y-M, Hsiao Y-Y, Fang L-S, et al. (2008) Cell cycle propagation is driven by light-dark stimulation in a cultured symbiotic dinoflagellate isolated from corals. Coral Reefs 27:823-835
51. Wijgerde T (2013) Coral Feeding: An Overview. Advanced Aquarist 12(12)
52. Wijgerde T, Laterveer M (2013) Coral growth under Light Emitting Diode and Light Emitting Plasma: a cross-family comparison. Advanced Aquarist 12(2)
53. Wijgerde T, Tilstra A (2014) Debunking Aquarium Myths. Advanced Aquarist 13(2)
54. Wijgerde T, van Melis A, Silva CIF, Leal MC, Vogels L, et al. (2014) Red Light Represses the Photophysiology of the Scleractinian Coral Stylophora pistillata. PLoS ONE 9(3): e92781. doi:10.1371/journal.pone.0092781
Inloggen
Registreren
Foto van de maand

Hippocampus erectus in het aquarium van David Hogenhuis
Foto: David Hogenhuis