Portaalsite voor de échte zeeaquariaan
Zoeken op de site
Partners








Dieren
Waar voeden koralen zich mee? Een overzicht
De idee dat koralen, net als alle andere dieren, moeten worden gevoed om goed te kunnen groeien, lijkt eindelijk de aquariumhobby te zijn doorgedrongen. Sinds de invloedrijke werken van koraalwetenschappers zoals Thomas Goreau en Leonard Muscatine, is onze kennis over hoe koralen aan voeding winnen gestaag toegenomen. Tegenwoordig weten we dat koralen die een mutualistische symbiose vormen met dinoflagellaten (Symbiodinium spp.) het grootste deel van hun koolstofenergie halen uit deze zogenaamde zoöxanthellen (zie ook Wijgerde 2013a). Koolstofrijke verbindingen zoals glucose en glycerol alleen zijn echter niet voldoende om koralen te laten groeien. Daarvoor zijn andere elementen zoals stikstof, fosfor en zwavel nodig. De huidige opvatting is dat zoöxanthellen hun gastheerkoraal niet van voldoende hoeveelheden van deze elementen voorzien zodat koralen volledig op de symbiose kunnen vertrouwen. Het is om deze reden dat koralen ook voeden door organische voedingsstoffen uit de externe omgeving op te nemen. Inderdaad, koralen voorzien van een externe, organische voedingsbron heeft uitgesproken effecten op hun groei. Dit roept belangrijke vragen op, zoals: waar voeden koralen zich mee, hoe wordt voeding beïnvloed door omgevingsfactoren en wat betekent dit voor de aquariumhobby en kostenefficiënte koraalaquacultuur?

Koralen kunnen zich voeden met voedseldeeltjes met mesenteriale filamenten, verlengingen van de maagwand. Hier zijn ze zichtbaar als witte strengen afkomstig van Stylophora pistillata-poliepen.
Waar voeden koralen zich mee?
Verplaatste voedingsstoffen van symbiotische zoöxanthellen, endolitische algen en stikstofbindende bacteriën.
Onder natuurlijke omstandigheden ontvangen koralen het grootste deel van hun organische koolstof van de endosymbiotische zoöxanthellen en in sommige gevallen van endolithische algen die zich in het koraalskelet bevinden (Muscatine et al. 1990; Fine en Loya 2002). Door gebruik te maken van lichtenergie zetten zoöxanthellen (genus Symbiodinium) en endolithische algen (genus Ostreobium) anorganische verbindingen verkregen uit het koraal en zeewater (kooldioxide, bicarbonaat, ammonium, nitraat, fosfaat) om in organische moleculen zoals glucose en glycerol. Dit proces staat bekend als fotosynthese of fotoautotrofie (van de Griekse woorden phos , of licht, autos , of zelf, en troph, of voeden), en het stelt de zoöxanthellen in staat zichzelf en hun gastheerkoraal te voeden, aangezien het overschot aan geproduceerde organische verbindingen wordt afgegeven aan de cellen van het koraal. De anorganische afvalproducten die door het koraal worden geproduceerd (kooldioxide, ammonium) worden vervolgens gerecycled door de zoöxanthellen. Bovendien voorzien stikstofbindende cyanobacteriën zoöxanthellen van ammoniak, die de bacteriën produceren uit opgeloste stikstof (N 2 ). Door deze uitwisseling van voedingsstoffen tussen koralen, zoöxanthellen en bacteriën kunnen koralen groeien in wat soms een woestijnachtige zee wordt genoemd, met een lage beschikbaarheid van voedingsstoffen (Muscatine 1990; Lesser et al. 2007).

Een beperking van fotosynthese is echter dat het koralen niet lijkt te voorzien van voldoende organische stikstof en fosfor om weefselgroei en organische matrixsynthese te behouden (zie hieronder). Daarom moeten koralen zich voeden met organisch materiaal, dat heterotrofie of heterotrofe voeding wordt genoemd (van de Griekse woorden heteros, of anders, en trof, of voeding). Hieronder zal ik de verschillende externe voedselbronnen beschrijven die koralen kunnen gebruiken. Het is belangrijk op te merken dat niet elke koraalsoort al deze bronnen kan gebruiken. Bovendien hangt de mate waarin koralen auto- of heterotroof zijn af van omgevingsomstandigheden, zoals de beschikbaarheid van licht en voedseldeeltjes.
Bacteriën en protisten
Hoewel het aannemelijk is dat koralen virussen opnemen en verteren (femtoplankton, deeltjesgrootte <0,2 µm), is vastgesteld dat koralen zich kunnen voeden met microben zoals (cyano)bacteriën en flagellaten. Deze microben worden geclassificeerd als picoplankton, met een deeltjesgrootte van 0,2 tot 2 µm, en nanoplankton, met een grootte van 2 tot 20 µm. Tijdens een studie over het voeden van koralen, Houlbrèque et al. (2004) geïncubeerd drie scleractinian koralen, Stylophora pistillata , Galaxea fascicularis en de azooxanthella Tubastraea aurea gedurende 6 uur in stroomkamers met pico- en nanoplankton (deeltjesgrootte <100 µm). Ze volgden veranderingen in de concentraties van bacteriën, cyanobacteriën en flagellaten tijdens de incubatie en ontdekten dat deze microben allemaal werden opgenomen door de drie koraalsoorten. Nanoflagellaten bleken een belangrijke bron van stikstof te zijn, wat belangrijk is voor de groei van koralen.
Sinds de jaren negentig is bekend dat koralen zich kunnen voeden met pelagische (vrij zwevende) microalgen, hoewel dit beperkt is tot octokoralen. De werken van Fabricius et al. (1995a,b) hebben aangetoond dat de koralen Dendronephthya, Nannochloropsis , Isochrysis en Tetraselmis Alcyonium dit doen (Elyakova et al. 1981).

Microscopische beelden van Stylophora pistillata-poliepen die Artemia-nauplii binnenkrijgen.
Microalgen (fytoplankton)
Sinds de jaren negentig is bekend dat koralen zich kunnen voeden met pelagische (vrij zwevende) microalgen, hoewel dit beperkt is tot octokoralen. De werken van Fabricius et al. (1995a,b) hebben aangetoond dat de koralen Dendronephthya hemprichi, D. sinaiensis, Scleronephthya corymbosa en Acabaria sp. voeden zich voornamelijk met fytoplankton, dat in het laboratorium Nannochloropsis, Isochrysis en Tetraselmis spp omvat. Het vermogen van octokoralen om zich te voeden met fytoplankton is waarschijnlijk gerelateerd aan de nauw uit elkaar geplaatste pinnulen op hun tentakels, evenals morfologische en gedragsaanpassingen aan het leven in sterke stroming (Fabricius 1995a). Deze bevinding komt goed overeen met het feit dat plantverterende carbohydrasen (amylase en laminarinase) zijn gevonden in koralen van het genus Alcyonium (Elyakova et al. 1981).
Op dit moment zijn er ook aanwijzingen dat scleractinische koralen zich kunnen voeden met microalgen, in tegenstelling tot eerdere overtuigingen. Tijdens een recente studie hebben Leal et al. (2013) ontdekt dat verschillende koralen zich konden voeden met fytoplankton, meer specifiek diatomeeën (Conticribra weissflogii , Thalassiosira pseudonana), een cryptofyt (Rhodomonas marina) en een haptofyt (Isochrysis galbana). Al deze algen worden beschouwd als nanoplankton, met een groottebereik van 4 tot 12 µm. Tijdens voedingsproeven werden zes koraalsoorten blootgesteld aan de verschillende algenstammen en na een uur werden de koralen gewassen met gefilterd zeewater en geanalyseerd op algen-DNA. De resultaten toonden aan dat het azooxanthellaat scleractinisch koraal Tubastraea coccinea zich kon voeden met C. weissflogii, T. pseudonana en I. galbana. Het zachte koraal Heteroxenia fuscecens werd gevoed met R. marina, het madreporaria koraal Pavona cactus nam R. marina en I. Galbana aan en het gematigde madreporaria koraal Oculina arbuscula werd gevoed met C. weissflogii en I. Galbana. Het scleractinisch koraal Stylophora pistillata en het zachte koraal Sinularia flexibilis leken zich met geen van de ge teste microalgen te voeden.
Deze resultaten geven aan dat elke koraalsoort een voorkeur heeft voor specifieke deeltjestypes, hoewel het nog niet duidelijk is hoe deze selectiviteit plaatsvindt. Ook moet nog worden onderzocht in hoeverre de koralen deze algen kunnen verteren, aangezien hun celwanden (kalkhoudende coccolieten of silica-frustules) het koraal nodig hebben om specifieke zuren en enzymen te produceren voor de spijsvertering. Osinga et al. (2012) rapporteerden het bestaan van borstelrand-enzymen in S. pistillata, wat suggereert dat scleractinische koralen plantaardig materiaal kunnen afbreken, hoewel plantverterend amylase en laminarinase begin jaren tachtig niet werden gevonden in steenkoralen (Elyakova et al. 1981). Bovendien toonden ze aan dat het koraal Pocillopora damicornis, een soort verwant aan S. pistillata (familie Pocilloporidae), een verhoogde groei vertoonde na enkele weken dagelijkse batchvoeding met de microalg Tetraselmis suecica . Ze vonden niet zo’n gunstig effect van Nannochloropsis sp., wat in overeenstemming lijkt te zijn met Leal et al. (2013), in die zin dat elke koraalsoort een specifieke voorkeur en verteringscapaciteit van bepaalde levende voedseldeeltjes kan hebben. Het vermogen van het koraal om bepaalde materialen te verteren kan verband houden met het microbiële consortium in de spijsverteringsholte van de poliep (coelenteron), waar verschillende aanwezige bacteriën de afbraak van specifieke componenten kunnen vergemakkelijken met behulp van spijsverteringsenzymen.
De reden waarom gevoede koralen verhoogde dichtheden van zoöxanthellen vertonen, is hoogstwaarschijnlijk dat verhoogde stikstofhoudende afvalproducten (zoals ammonium/NH4 +) die door het koraal worden uitgescheiden, de groei van zoöxanthellen bevorderen. Op zijn beurt kan voeding de zoöxanthellen ook overhalen om meer aminozuren te produceren en naar hun koraalgastheer te verplaatsen, wat de groei van zacht weefsel en organische matrixsynthese ten goede komt (Swanson en Hoegh-Guldberg 1998; Wang en Douglas 1999).
Benthische (macro)algen
In verband met het voeden met microalgen, kunnen koralen zich ook voeden met bentische algen. Tijdens een koraalverbleking in 2011 bleken de steenkoralen Colpophyllia natans en Montastraea faveolata zich te voeden met algengras en Dictyota macroalgen (Marhaver 2011). De koralen strekten hun mesenteriale filamenten (uitsteeksels van de gastroderm of maagwand) uit door de mond of zijkanten van de poliepen en kwamen fysiek in contact met verschillende soorten algen. Het is bekend dat deze mesenteriale of maagfilamenten cnidocyten en spijsverteringscellen bevatten, waardoor koralen naburige koralen kunnen doden en verteren, en hun rol bij het verkrijgen van voedingsstoffen wordt steeds duidelijker (Wijgerde et al. 2011). Door enzymen af te scheiden, zorgen mesenteriale filamenten ervoor dat koralen voedseldeeltjes extern kunnen verteren, waarna gespecialiseerde cellen in de filamenten de vrijgekomen voedingsstoffen kunnen opnemen. Door zich te voeden met bentische algen, of hun koolstofrijke uitscheidingen, compenseerden de gebleekte koralen mogelijk het verlies van voedingsstoffen die normaal via hun zoöxanthellen worden gewonnen. Marhaver (2011).
Een soortgelijke waarneming werd gedaan in ons laboratorium aan de Wageningen Universiteit, waar verschillende S. pistillata-kolonies een uitgesproken uitdrijving van mesenteriale filamenten vertoonden in gebieden waar een biofilm zich ophoopte door stilstaand water. Omdat deze biofilms rijk zijn aan organische verbindingen en bacteriën, kan dit verklaren waarom de koralen deze gebieden met filamenten hebben bezaaid.
Zeegrassen
Een laatste studie die suggereert dat koralen zich kunnen voeden met (lagere of hogere) planten werd uitgevoerd door Lai et al. (2013), die ontdekte dat het scleractinische koraal Oulastrea crispata zeegras deeltjes opneemt (Halophila ovalis). Ze stelden koraalkolonies gedurende twee uur bloot aan zeegrasdeeltjes die waren gelabeld met een stabiele stikstofisotoop ( 15 N), waarna ze het coelenterische gehalte van tien poliepen uit elke kolonie extraheren. Toen een kolonie positief bleek te zijn voor zeegras, hebben ze het weefsel schoongemaakt, verwijderd en geanalyseerd op 15 N. Hieruit bleek dat O. crispata Halophila ovalis opneemt in deeltjes, en mogelijk organische voedingsstoffen die uit dit zeegras worden verkregen, verteert en assimileert. Ze ontdekten ook dat dit koraal organische verbindingen uit het zeegras absorbeerde. Deze studie suggereert dat koralen die in de buurt van zeegrasweiden leven, zich kunnen voeden met losgeraakt zeegrasmateriaal en zeegras-exsudaten. Net als bij de studie van Leal et al. (2013), moet nog worden bepaald in welke mate koralen zeegrasmateriaal kunnen verteren en assimileren.
Zoöplankton
Het vermogen van koralen om zich te voeden met zoöplankton is in de loop der jaren uitgebreid bestudeerd, vooral wanneer rekening wordt gehouden met scleractinische koralen (Houlbrèque en Ferrier-Pagès 2009; Ferrier-Pagès et al. 2011).
Over het algemeen zijn deze koralen zeer goed in staat om zoöplankton te vangen dankzij hun krachtige cnidocyten, die capsules bevatten die vol zitten met neurotoxinen en met lasso-achtige strengen kunnen vangen. Bovendien gebruiken deze koralen slijm om levende prooien te vangen. Door gebruik te maken van microscopisch kleine haartjes, cilia genaamd, transporteren de poliepen van het koraal prooi-items (in) naar de mond. Zoöplankton kan ook extern worden verteerd met mesenteriale filamenten.

Copepoden vormen een belangrijke voedselbron voor scleractinian koralen.
Op koraalriffen omvat het zoöplankton dat door steenkoralen wordt belaagd schaaldieren zoals roeipootkreeftjes, vlokreeften, ostracoden, mysiden, wormen zoals polychaeten en chaetognaths (pijlwormen) en veel dierlijke larven. In het aquarium zijn veel van deze natuurlijke prooi-items niet beschikbaar, en levende of dode Artemia en Mysis zijn veelvoorkomende feeds. Onderzoek heeft aangetoond dat Artemia-nauplii, hoewel ze van nature niet beschikbaar zijn voor koralen, een zeer geschikte voeding zijn, die de koraalgroei aanzienlijk verbetert (zie ook hieronder).
Zwarte koralen hebben, als verwanten van scleractinische koralen, ook het vermogen om zoöplankton te vangen en te verlammen. Laboratoriumexperimenten op Antipathes grandis tonen aan dat zijn poliepen amfipoden, roeipootkreeften en chaetognaths kunnen vangen. Net als bij steenkoralen vindt vangst plaats door het gebruik van tentakels en slijm, waarna trilhaartjes op het ectoderm voedsel naar de mond transporteren (Bo 2009). Zwarte koralen met grote poliepen, zoals die van Antipathes en Cirrhipathes spp., kunnen roeipootkreeftjes van minstens 1700 m groot opnemen.
Octokoralen variëren in hun vermogen om zoöplankton te vangen en vast te houden, en zachte koralen lijken specifiek minder aangepast aan deze klasse van prooien. Zachte koralen van de genera Sinularia, Sarcophyton, Cladiella, Nephthea, Dendronephthya en Paralemnalia zijn bijvoorbeeld niet in staat groter zoöplankton vast te houden na vangst (Fabricius et al. 1995a). Bijvoorbeeld Dendronephthya hemprichi vangt alleen klein en zwak zwemmend zoöplankton, zoals tweekleppige en buikpotige larven, ostracoden, amfipoden, tintinniden (ciliaten), polychaeten en viseieren. Items kleiner dan 300 µm worden gevangen en binnen 10 tot 20 seconden opgenomen, maar prooien van 750 µm groot en groter worden nauwelijks gevangen en ontsnappen meestal binnen een minuut.
Interessant is dat wanneer Dendronephthya spp. grotere zoöplankton vangen, er geen tekenen van verlamming zijn, zelfs niet na enkele minuten, of wanneer deze deeltjes meerdere keren worden gevangen. Het lijkt erop dat Dendronephthya spp. en andere zachte koralen niet beschikken over voldoende ontwikkelde cnidocyten om grotere prooien effectief te verlammen. Er is inderdaad gevonden dat de nematocysten-gifcapsules in de cnidocyten van veel octocoralen slecht ontwikkeld zijn (Fabricius en Alderslade 2001).

Gorgonische octokoralen zijn echte alleseters en kunnen zich voeden met zoöplankton, fytoplankton en afval.
Vergelijkbare resultaten zijn gevonden met octocoralen zoals Dendronephthya spp. en de mediterrane gorgonen Paramuricea clavata en Corallium rubrum, die zich ook voeden met bacteriën en protisten (Fabricius et al. 1995a,b; Picciano en Ferrier-Pagès 2007; Ribes et al. 1999). Hoewel bacteriën slechts een klein deel van de totale koolstoftoevoer uitmaken, kunnen ze een belangrijke bron van stikstof zijn.
Gorgonen zijn octokoralen die over het algemeen goed zijn aangepast om zoöplankton te vangen. Het is bekend dat verschillende gorgonen, waaronder de tropische soorten Subergorgia suberosa, Melithaea ochracea en Acanthogorgia vegae, actief zwemmende Artemia-nauplii in het laboratorium kunnen vangen (Dai en Lin 1993; Lin et al. 2002).
Hydrozoaire koralen (familie Milleporidae, of vuurkoralen, en familie Stylasteridae, of kantkoralen) dragen ten slotte krachtige cnidocyten op hun tentakels waardoor ze zoöplankton efficiënt kunnen vangen. Het is inderdaad bekend dat deze koralenvraatzuchtige zoöplanktonvoeders zijn (Lewis 2006). Hun nematocysten vuren met zo’n kracht dat zelfs de menselijke huid er gevoelig voor is, het aanraken van deze koralen veroorzaakt een intens branderig gevoel en huiduitslag. In tegenstelling tot andere koralen, wordt verlamming en opname van prooien opgevangen door twee soorten poliepen. Prooien worden gestoken door verdedigende stekende poliepen die dactylozooiden worden genoemd, terwijl prooien worden ingenomen en verteerd door gastrozooiden. Elke gastrozooid is omgeven door vijf tot vijftien dactylozooiden, de laatste veel langer en dunner.
Vissen
Koralen met grotere poliepen kunnen kleine vissen heel verslinden, wat soms in het aquarium wordt waargenomen. Dit kunnen vissen zijn die om de een of andere reden verzwakt zijn en als gevolg daarvan weerloos worden overgelaten aan de tentakels en cnidocyten van grote koralen. Scolymia spp., Fungia spp. en Trachyphyllia geoffroyi zijn voorbeelden van koralen die dit gedrag vertonen.
Andere koralen
Een interessante voedselbron voor koralen zijn de koralen zelf. Op het rif is gevonden dat koralen zich voeden met naburige kolonies, die ze aanvallen en extern verteren met mesenteriale filamenten. Dit gedrag kan in de eerste plaats een vorm van effectieve competitie zijn tussen soorten en individuen binnen soorten (respectievelijk de zogenaamde interspecifieke en intraspecifieke competitie), maar het biedt koralen ook een extra voedselbron.

Een Millepora sp., mogelijk M. dichotoma, met zijn krachtige dactylozooids uitgebreid (Rode Zee, Egypte).
Afval
Detritus is een verzamelnaam voor organische deeltjes die ontstaan uit ontlasting, voedselresten en rottende organismen. Afvalstoffen komen veel voor op koraalriffen en in het aquarium en bezinken langzaam op de bodem als sediment. Dit sediment bevat bacteriën, protozoa, microscopisch kleine ongewervelde dieren, microalgen en organisch materiaal. Deze sedimentaire bronnen kunnen allemaal dienen als koraalvoedingsstoffen wanneer ze worden gesuspendeerd, vooral voor soorten die in troebel water groeien. Experimenten hebben aangetoond dat veel scleractinische koralen afval kunnen opnemen en assimileren (bijv. Anthony 1999,2000; Anthony en Fabricius 2000; Roff et al. 2009), dat vastzit in koraalslijm.
Hoewel steenkoralen afval kunnen opnemen als het beschikbaar is, is gebleken dat verschillende gorgonen zich voornamelijk voeden met zwevend afval. Bijvoorbeeld, de mediterrane gorgonen Corallium rubrum, Paramuricea clavata en Leptogorgia sarmentosa verwerven het grootste deel van hun koolstof als afval (Ribes et al. 1999; Tsounis et al. 2006). Dit lijkt ook te gelden voor sommige van hun tropische tegenhangers, zoals Menella en Swiftia spp. Deze gorgonen vangen en nemen gemakkelijk kleine gepelleteerde visvoeders in het aquarium op.

Een kleine Galaxea fascicularis-kolonie die afvalstof in slijmnetten heeft gevangen.
https://youtu.be/y0hm3ads15w
Link naar een video van een gorgonen (Menella sp.) die zich voedt met deeltjes organisch materiaal van 5-800 µm groot (droog visvoer). Hoewel het technisch gezien geen afval is, is droog visvoer vergelijkbaar omdat het niet-levend organisch materiaal is dat wordt geproduceerd uit dieren en planten.
Koralen die in diepere wateren leven, gebruiken ook afvalstof als een belangrijke voedingsbron. Het diepwater scleractinisch koraal Lophelia pertusa (maar ook gorgonen en zwarte koralen) vangt zeesneeuw op, of afvalmateriaal dat via neerwaartse stromingen vanuit hogere oceanische lagen naar de diepte wordt gebracht (Bo 2009; Davies et al. 2009). Er moet echter worden opgemerkt dat te veel sediment schadelijk is voor koralen en riffen. Hoge sedimentatie verstikt het rif letterlijk door licht, voeding en gasuitwisseling te blokkeren (Erftemeijer et al. 2012).
Opgelost organisch materiaal
Opgeloste organische stof (DOM) is een belangrijke voedselbron voor veel koralen. Hoewel bekend is dat koralen organisch materiaal afscheiden, voornamelijk via slijmafgifte, nemen ze wel opgeloste organische stoffen op uit het water. Met radioactieve tracers werd ontdekt dat scleractinische koralen opgeloste glucose uit het water opnemen. Meer ecologisch relevant, koralen kunnen ook aminozuren en ureum uit het zeewater opnemen (Grover et al. 2006, 2008). Hoewel deze stoffen slechts in minieme concentraties op koraalriffen aanwezig zijn, vormen ze een belangrijke bron van organische stikstof. Voor Stylophora pistillata, kan de opname van aminozuren 21% van het stikstofbudget uitmaken (Grover et al. 2008), hoewel de balans tussen deze opname en de opname van voedingsstoffen uit andere bronnen afhangt van wat beschikbaar is voor het koraal. Aminozuren zijn belangrijk voor de synthese van de organische matrix, een extracellulair eiwitachtig raamwerk dat nodig is voor skeletgroei in koralen (zie hieronder). Het is intrigerend dat koralen ook ureum uit het water opnemen. Dit geeft aan dat koralen zich mogelijk hebben aangepast aan de aanwezigheid van hogere dieren op het rif, zoals vissen, die gezamenlijk dagelijks grote hoeveelheden van deze stikstofverbinding produceren.
Koralen nemen niet alleen organische stoffen op, ze lijken deze ook in het water op te sporen. Een veel voorkomende observatie is de verlenging van koraaltentakels na toevoeging van plankton of organische stoffen aan het aquariumwater. Toevoeging van de aminozuren glycine, alanine of glutamaat aan het water resulteert in tentakelverlenging, zwelling van weefsel (coenenchym) en soms extrusie van mesenteriale filamenten (Goreau et al. 1971). Net zoals de menselijke tong receptoren heeft om veel stoffen te detecteren, kunnen koralen ook receptoren hebben ontwikkeld die organische verbindingen zoals aminozuren herkennen. Het vermogen om aminozuren in het water te detecteren, kan dienen om zoöplankton waar te nemen, waardoor koralen zich kunnen voorbereiden op het vangen van prooien.
Voedselbronnen die door koralen worden gebruikt voor de opname van energie en voedingsstoffen kunnen worden onderverdeeld in interne en externe bronnen. Interne bronnen zijn stikstofbindende bacteriën, die opgeloste stikstof (N2) omzetten in ammoniak (NH3), een proces dat diazotrofie wordt genoemd, en zoöxanthellen, die de ammoniak omzetten in aminozuren en eiwitten. Daarnaast zetten zoöxanthellen kooldioxide (CO2) tot glycerol, glucose, vetzuren en aminozuren via een proces dat bekend staat als fotosynthese, een vorm van autotrofie. Een groot deel van deze organische verbindingen wordt verplaatst naar de gastheercellen van het koraal, die deze voornamelijk gebruiken om in hun energiebehoefte te voorzien. Externe bronnen zijn fijn stof en opgeloste organische stof, die uit de waterkolom worden opgenomen. Koralen voeden zich met fytoplankton en bentische algen (herbivoor), zoöplankton, kleine vissen en andere koralen (carnivoor), bacteriën en protisten (microheterotrofie), zwevende deeltjes (detrivory) en tenslotte opgeloste organische stoffen zoals ureum en aminozuren. Deze opname van deeltjes en opgelost organisch materiaal uit de waterkolom staat bekend als heterotrofie, en de organische verbindingen die uit dit proces worden verkregen, worden door het koraal gebruikt voor energieproductie en groei. Om tot nu toe te concluderen, is het duidelijk dat koralen in staat zijn om organische verbindingen uit een breed scala van bronnen op te nemen, wat de diverse en efficiënte aard van koralen als alleseters onderstreept.
Opgelost anorganisch materiaal
Hoewel dit artikel zich richt op het voeden met organische verbindingen, nemen koralen ook anorganische stoffen op uit de waterkolom. Ik zal kort de belangrijkste elementen noemen die in anorganische vorm zijn opgenomen. Deze omvatten, maar zijn niet beperkt tot, anorganische stikstof (opgeloste stikstof/N2 , ammonium/NH4+ en nitraat/NO3–) en fosfor (fosfaat, HPO42-), anorganische koolstof (kooldioxide/CO2, bicarbonaat/HCO3–), alkalimetalen (natrium/Na+, kalium/K+), aardalkalimetalen (calcium/Ca2+, magnesium/Mg2+, strontium/Sr2+), overgangsmetalen (bijv. zink/Zn2+, ijzer/Fe2/3+, koper/Cu2+, mangaan/Mn2+), metalloïden (Boor/B) en niet-metalen (jodium als jodide/I– en jodaat/IO3–, zuurstof/O2). De opname van anorganische stikstof en fosfor is te wijten aan de aanwezigheid van symbiotische zoöxanthellen en bacteriën, die deze omzetten in organische verbindingen voor hun groei.
De opname van niet-metaalverbindingen is te danken aan zowel de koraalgastheer als zijn symbionten. Calcium en magnesium zijn belangrijk voor koraalverkalking, koolstofdioxide en bicarbonaat zijn essentieel voor fotosynthese en koraalverkalking, sporenelementen zoals zink en jodium worden door koralen en symbionten gebruikt voor de enzymfunctie en mogelijk hormoonproductie, en zuurstof is belangrijk voor de ademhaling.

Effecten van voeding op koraalgroei en fysiologie
De effecten van voeding op de groei en fysiologie van koralen zijn goed bestudeerd en worden uitgebreid besproken in recente beoordelingen door Houlbrèque en Ferrier-Pagès (2009) en Ferrier-Pagès et al. (2011). Tot nu toe hebben de meeste studies zich gericht op de effecten van zoöplanktonvoeding, voornamelijk Artemia-nauplii, op de groei en fysiologie van koralen.
Fotosynthese en zoöxanthellendichtheid
Onderzoek heeft aangetoond dat voeding de fotosynthesesnelheid van zoöxanthellenkoralen verbetert, door de dichtheid van zoöxanthellen en het chlorofylgehalte te verhogen. Voor S. pistillata verdubbelt de zooxanthellae-dichtheid binnen enkele weken na zoöplanktonvoeding, zowel bij lage als hoge lichtniveaus. Het aantal dinoflagellaten dat zich in een enkele koraalgastcel bevindt, neemt ook toe, met maximaal vier zoöxanthellen per koraalcel. Een hogere fotosynthesecapaciteit stelt het koraal in staat meer lichtenergie om te zetten in chemische energie, die kan worden gebruikt voor groei.
De reden waarom gevoede koralen verhoogde dichtheden van zoöxanthellen vertonen, is hoogstwaarschijnlijk dat verhoogde stikstofhoudende afvalproducten (zoals ammonium/NH4+) die door het koraal worden uitgescheiden, de groei van zoöxanthellen bevorderen. Op zijn beurt kan voeding de zoöxanthellen ook overhalen om meer aminozuren te produceren en naar hun koraalgastheer te verplaatsen, wat de groei van zacht weefsel en de organische matrixsynthese ten goede komt (Swanson en Hoegh-Guldberg 1998; Wang en Douglas 1999).
Verkalking en de organische matrix
Naast het stimuleren van de fotosynthese, verhoogt het voeren de verkalkingssnelheid in zoöxanthellen-scleractinische koralen. Na acht weken voeden met zoöplankton ( Artemia-nauplii), verdubbelt de verkalkingssnelheid van Stylophora pistillata. Verschillende mechanismen kunnen verantwoordelijk zijn voor dit fenomeen. Ten eerste kan voeren verkalking stimuleren via een verhoogde bicarbonaatproductie. Voeren verhoogt de massa van koraalweefsel en daarmee de productie van metabolisch CO2 . Een deel van deze CO2 wordt enzymatisch omgezet in bicarbonaat, dat als substraat voor verkalking kan worden gebruikt. Voor het koraal Stylophora pistillata, is berekend dat het ongeveer 75% van zijn bicarbonaat uit zijn eigen metabolisme kan verkrijgen. Ten tweede zorgt meer voeding voor meer chemische energie, direct maar ook indirect door de fotosynthetische capaciteit van het koraal te vergroten (zie hierboven), waardoor er meer calciumionen naar het groeiende skelet kunnen worden getransporteerd. Ten slotte kan voeding verkalking stimuleren door de synthese van de organische matrix te verbeteren via een verhoogde toevoer van aminozuren.
De organische matrix is een extracellulair eiwitraamwerk dat wordt uitgescheiden door koraalcellen en essentieel is voor de vorming van skeletten. Het biedt een kiemplaats voor aragoniet (calciumcarbonaat) kristallen om te groeien, en stimuleert en reguleert hun vorming (Allemand et al. 1998, 2004). Omdat de organische matrix rijk is aan aminozuren zoals asparaginezuur, kan voeding de synthese van de organische matrix en dus verkalking verbeteren door de toevoer van dit aminozuur te vergroten.
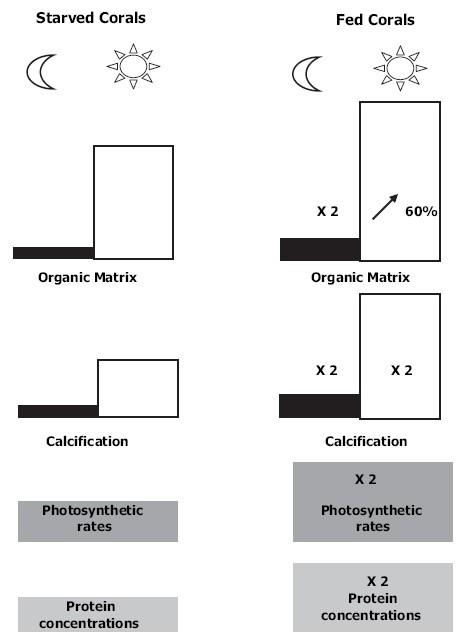
Opgemerkt moet worden dat zware voeding van koralen nadelen kan hebben. In ons laboratorium van Wageningen UR hebben we de korte termijn effecten bestudeerd van het voeden van zoöplankton op de lichte en donkere verkalking van het koraal Galaxea fascicularis. Hoewel voeding weinig effect had op de groei onder lichte omstandigheden, waren de verkalkingspercentages van gevoede koralen in totale duisternis bijna nul (Wijgerde et al. 2012b). Onze huidige hypothese is dat donkere verkalking wordt geremd door zware voeding als gevolg van een tijdelijke acidose van koraalweefsel, veroorzaakt door verhoogde metabolische activiteit. Tijdens de nachtelijke voeding kunnen koralen energie investeren in de groei van zacht weefsel en de synthese van organische matrix, in plaats van verkalking.

Enkele voorbeelden van vraatzuchtige koralen met uitgestrekte tentakels. Linksboven: Acanthastrea lordhowensis. Rechtsboven: Caulastraea sp. Linksonder: Tubastraea sp. Rechtsonder: Trachyphyllia geoffroyi.
Koraal weefsel
Naast het verbeteren van de fotosynthese, de dichtheid van zoöxanthellen, de calcificatiesnelheid en de organische matrixsynthese, verhoogt het voeren van zoöplankton ook het eiwit- en vetgehalte van zacht weefsel. Verzadigde en onverzadigde vetzuren, evenals alcoholen en sterolen nemen in concentratie toe na langdurige voeding met Artemia-nauplii. Door de toename van lipidenvoorraden kunnen koralen beter omgaan met stress, vooral bleken. Wanneer hoge watertemperaturen een verlies van zoöxanthellen veroorzaken, kunnen koralen niet langer fotosynthese uitvoeren en moeten ze vertrouwen op het vangen van prooien en energiereserves om te overleven (Grottoli et al. 2006).
Wat bepaalt de voedingssnelheid van koraal?
Waterstroom
Waterstroming is om veel verschillende redenen essentieel voor koralen. Naast het verbeteren van de gasuitwisseling en het bevorderen van de verwijdering van sediment, zorgt de waterstroom ervoor dat koralen zich kunnen voeden met (levende) deeltjes (Wijgerde 2013 en referenties daarin). Omdat de belangrijke rol van waterbeweging vrij duidelijk is, is waterstroming een van de best bestudeerde factoren die het vangen van prooien door koralen beïnvloeden.
Waterstroom heeft zowel gunstige als nadelige effecten op de koraalvoeding, afhankelijk van de stroomsnelheid. Hogere stroomsnelheden zullen de instroom van voedseldeeltjes vergroten en zullen daarom ten goede komen aan het voeren. Hogere stroomsnelheden zullen echter ook de kinetische energie van voedseldeeltjes verhogen, wat het vangvermogen van koraalpoliepen zal beperken. Bovendien zorgt een sterke waterstroom voor weerstandskrachten, wat resulteert in vervormde poliepen en een kleiner vangoppervlak en minder efficiëntie.
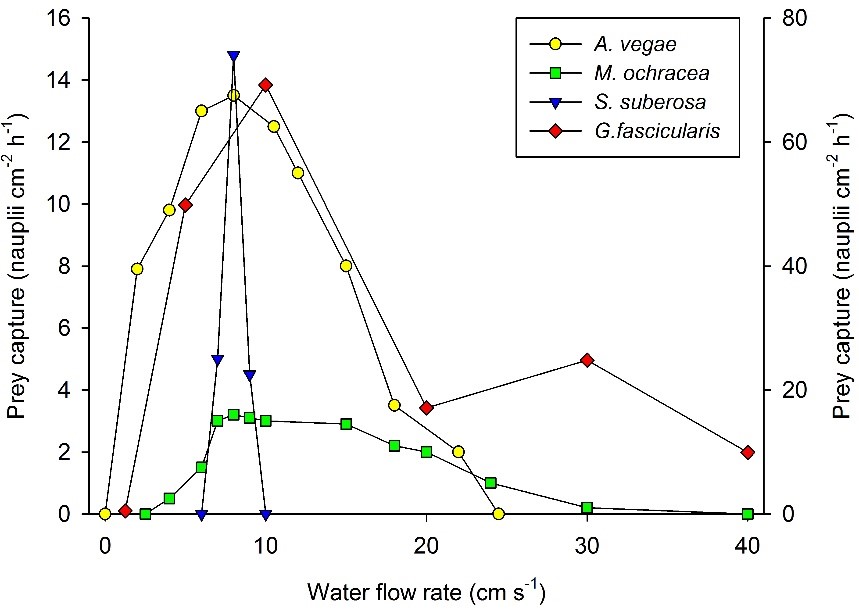
Relatie tussen waterstroomsnelheid en prooivangst voor vier koraalsoorten. Linker as: Acanthogorgia vegae, Melithaea ochracea en Subergorgia suberosa (prooiconcentratie: 20 individuen L-1). Rechter as: Galaxea fascicularis (prooiconcentratie: 10.000 nauplii L-1). Waarden zijn gemiddelden (N=2-4). Voor de duidelijkheid zijn de standaarddeviaties weggelaten. Na Dai en Lin (1993) en Wijgerde et al. (2012d).
Deze mechanismen verklaren waarom voor verschillende koraalsoorten klokvormige relaties tussen waterstroomsnelheid en prooivangst zijn gevonden, waarbij optima vaak tussen een stroombereik van 5 tot 10 cm/s-1 ligt . De bovenstaande grafiek illustreert hoe stroming de voedingssnelheid van vier verschillende koralen beïnvloedt; de octokoralen Acanthogorgia vegae, Melithaea ochracea en Subergorgia suberosa , en het scleractinische koraal Galaxea fascicularis . De grafiek toont direct de soortspecifieke reactie op stroming. De verschillende manieren waarop deze soorten reageren op waterstroming in termen van prooivangst, kunnen worden verklaard door verschillen in poliepmorfologie (zie hieronder).
Koraal maat
Naast de waterstroomsnelheid heeft de koloniegrootte invloed op heterotrofe voeding. De grootte van kolonies kan de voedingssnelheid van individuele poliepen beïnvloeden, zowel in negatieve als positieve zin, als gevolg van poliepinteracties binnen kolonies. Negatieve effecten zijn onder meer poliepschaduw (dwz poliepen die elkaar bedekken en belemmeren) en lokale uitputting van deeltjes, wat resulteert in verminderde prooivangst door stroomafwaartse poliepen (Hunter 1989). Positieve effecten zijn onder meer het genereren van intrakoloniale turbulentie en slijmafscheiding door stroomopwaartse poliepen, waardoor het vangen van prooien door stroomafwaartse poliepen wordt verbeterd (Wijgerde 2013 en referenties daarin).
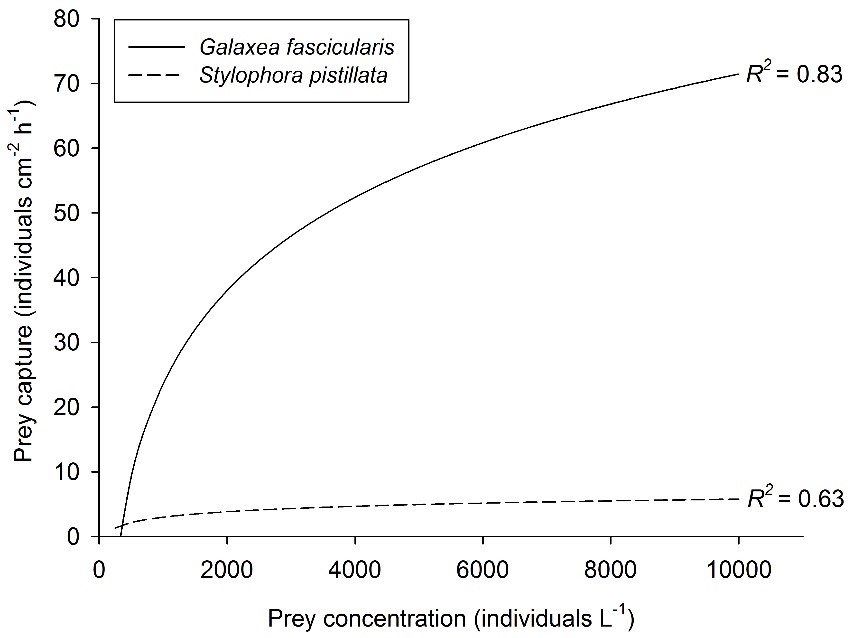
Verband tussen prooiconcentratie en prooivangst voor twee koraalsoorten, Galaxea fascicularis (Artemia nauplii) en Stylophora pistillata (Mediterraan zoöplankton). Best passende curven, respectievelijk N=30 en N=25. Nadat Ferrier-Pagès et al. (2003) en Wijgerde et al. (2011a, 2012c).
Hoewel poliepen in kolonies hogere voersnelheden kunnen vertonen in vergelijking met solitaire poliepen, lijkt de kolonie als geheel minder efficiënt te worden. In ons laboratorium ontdekten we dat slechts 7,7% van de poliepen in een kleine Galaxea fascicularis zoöplankton vangen. Dit betekent dat deze koralen per poliep minder voedsel vangen dan individuele poliepen. Deze waarneming past goed bij de afname van de relatieve groei van Galaxea met toenemende omvang (Schutter et al. 2010; Wijgerde et al. 2012a). Na 245 dagen incubatie vertonen kleine Galaxea-kolonies een reductie van 76% (van 2,5 tot 0,6% dag -1 ) in relatieve groei in vergelijking met enkele poliepen.
Prooi concentratie
Prooiconcentratie is een goed bestudeerde factor die de voedingssnelheid van koraal beïnvloedt. Hogere prooiconcentraties zullen de prooiaankomst van koraalpoliepen verhogen, wat een positief effect heeft op de voedingssnelheid. In eerste instantie wordt een lineair verband gevonden tussen prooidichtheid en koraalvoersnelheid. Wanneer de prooiconcentratie echter hoog genoeg wordt, wordt een verzadigend effect waargenomen. Dit komt omdat koraalpoliepen zijn gebonden aan een maximum aantal prooien dat ze op een bepaald moment kunnen vangen, opnemen en verteren. Dit verzadigende effect van prooiconcentratie wordt geïllustreerd door de onderstaande grafiek, die stabiliserende voedingssnelheden van de koralen Galaxea fascicularis en Stylophora pistillata laat zien met toenemende beschikbaarheid van prooien.

Rhizotrochus typus is een soort met grote, solitaire poliepen die vissen en garnalen kunnen verslinden.
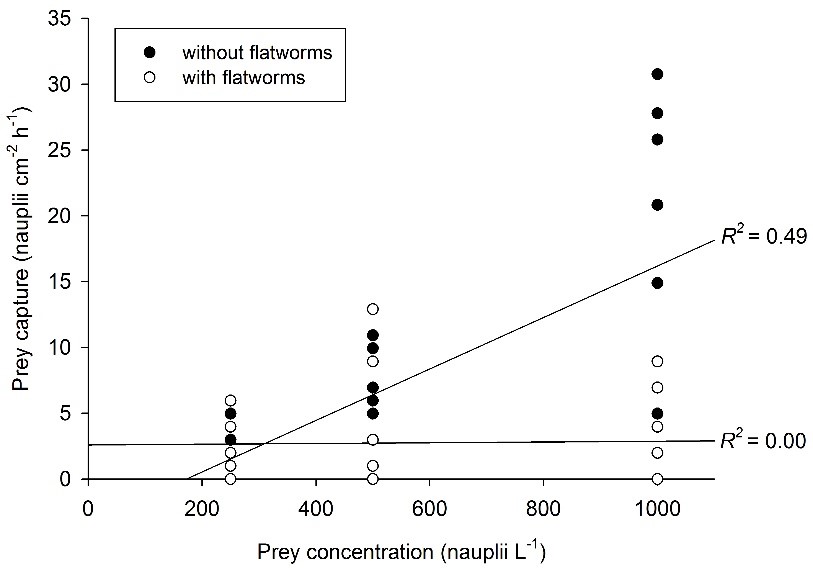
Relatie tussen prooiconcentratie en prooivangst voor Galaxea fascicularis poliepen, met (N=27) en zonder (N=27) epizoïsche platwormen. Een significante, positieve correlatie tussen prooiconcentratie en prooivangst wordt alleen gevonden voor wormvrije poliepen. Nadat Wijgerde et al. (2012c).
Poliep morfologie
De morfologie van koraalpoliepen is nog een andere factor die de koraalvoeding kan beïnvloeden. Zo heeft Subergorgia suberosa grote poliepen en stuit hoge trekkrachten, waardoor sterke stromingen haar poliepen gemakkelijk vervormen. Dit helpt verklaren waarom deze soort zich voedt in een smal bereik van stroomsnelheden, zoals te zien is in de bovenstaande stroomgrafiek. Melithaea ochracea daarentegen heeft kleinere poliepen en ondervindt daarom een lagere weerstand, wat resulteert in minder poliepvervorming bij sterkere stromingen. Dit is waarschijnlijk de reden waarom het zich in een breder scala aan stroomsnelheden voedt.
Hoewel ze gemakkelijker vervormd kunnen worden, kunnen grotere poliepen een hogere voedingscapaciteit hebben. Dit is duidelijk te zien in de bovenstaande grafiek, waar de grotere Galaxea fascicularis-poliepen (~5 mm corallietdiameter) significant meer prooien vangen in vergelijking met de veel kleinere Stylophora pistillata-poliepen (~1 mm corallietdiameter). Dit is waarschijnlijk te wijten aan het feit dat de poliepen van G. fascicularis grote hoeveelheden prooien extern kunnen verteren.
Een ander resultaat van poliepafmetingen is de maximale grootte die prooikoralen kunnen opnemen. Soorten met kleine poliepen kunnen roeipootkreeftjes en verschillende dierlijke larven vangen als maximale prooigrootte, terwijl soorten met grote poliepen (bijv. Fungiidae, Mussidae en Flabellidae) grote prooidieren zoals vissen en garnalen kunnen consumeren.
Epizoïsche platwormen
De laatste factor waar ik het hier over wil hebben, is de aanwezigheid van epizoïsche acoelomorfe platwormen. Deze platwormen worden in de aquariumhobby gewoonlijk planaria genoemd, hoewel dit eigenlijk wormen zijn van de geslachten Waminoa en Convolutriloba. Er is gesuggereerd dat deze wormen koralen negatief kunnen beïnvloeden door de hoeveelheid beschikbaar licht voor het koraal te verminderen en door de koraalslijmlaag te verwijderen (Barneah et al. 2007; Naumann et al. 2010). Bovendien concurreren Waminoa met hun gastkoraal Galaxea fascicularis om plankton, beperken ze de voedingscapaciteit van dit koraal aanzienlijk en stelen ze prooien van hun gastkoraal (Wijgerde et al. 2011b, 2012c).
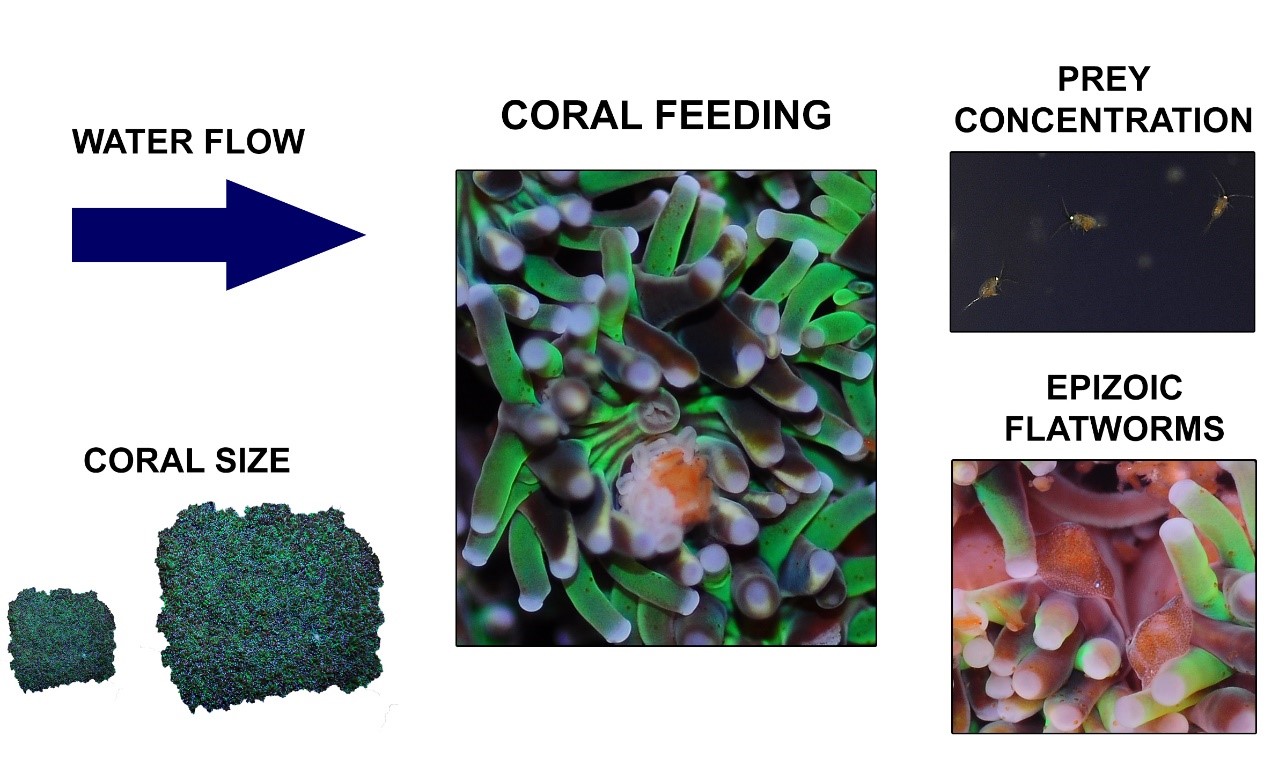
Overzicht van de verschillende factoren waarvan bekend is dat ze de voedingssnelheid van Galaxea fascicularis bepalen, waaronder waterstroom, koraalgrootte, prooiconcentratie en epizoïsche platwormen.
Bij het verstrekken van prooi aan G. fascicularis wordt alleen bij wormvrije poliepen een positief effect van prooiconcentratie op de voedersnelheid gevonden. Wanneer platwormen in hoge dichtheden aanwezig zijn (~ 3 tot 4 platwormen per poliep), zijn de voedingssnelheden van G. fascicularis beperkt tot ongeveer 2,5 prooi cm -2 h -1 . Op basis van deze bevindingen kunnen epizoïsche platwormen beter worden geclassificeerd als parasieten dan als commensalen, omdat hun aanwezigheid de groei en gezondheid van koralen negatief kan beïnvloeden. Uit veldgegevens blijkt inderdaad dat platwormen ernstige weefselnecrose in koralen veroorzaken (Hoeksema en Farenzena 2012).
Het is duidelijk dat de effecten van waterstroom, koraalgrootte, prooiconcentratie, poliepmorfologie en platwormen belangrijke implicaties hebben voor koraalaquacultuur en de aquariumhobby. Hieronder zal ik bespreken hoe deze kennis kan worden gebruikt om de voedingssnelheid van koralen en dus de groei te maximaliseren.
Koralen voeren in gevangenschap: maximale voedingssnelheid en groei
Gewapend met de hierboven beschreven kennis, kunnen we proberen de voedingssnelheid en groei van koraal te maximaliseren door de aquariumomgeving te manipuleren. Hieronder worden een aantal belangrijke strategieën beschreven die de aquariumliefhebber hierbij kunnen helpen.
Optimaal waterdebiet
Zoals hierboven vermeld, is waterstroming essentieel voor koralen, wat de groei, fotosynthese, gas- en warmteuitwisseling en sedimentverwijdering bevordert (Mass et al. 2010; Schutter et al. 2010, 2011; Jimenez et al. 2011; Erftemeijer et al. 2012 ). Hoewel veel koralen met variabele stroomsnelheden zullen groeien, kan elke soort (en genotype binnen soorten) optimaal groeien met een specifiek stroombereik. Dit is, althans gedeeltelijk, te wijten aan optimale voedingssnelheden binnen een bepaald stroombereik. De koralen Acanthogorgia vegae, Melithaea ochracea en Subergorgia suberosa vangen bijvoorbeeld allemaal zoöplankton optimaal op bij een stroomsnelheid van 8 cm/s -1 , mogelijk een weerspiegeling van hun gedeelde habitat. Bovendien vangt Subbergorgia suberosa alleen voedsel bij een zeer smal stroombereik, waardoor het onderhoud van deze soort zeer moeilijk is. Herbivore octokoralen hebben mogelijk meer stroming nodig voor een optimale deeltjesvangst. Het octokoraal Dendronephthya hemprichi, dat zich voornamelijk voedt met fytoplankton, voedt en groeit optimaal binnen een stroombereik van 10 tot 25 cm s- 1 (Fabricius et al. 2005a).
Dendronephthya en Scleronephthya spp. produceren dicht opeengepakte sklerieten die koloniestabiliteit bieden onder sterke waterstroom.
Tabel 1 geeft een overzicht van de optimale stroomsnelheden in termen van deeltjesvangst voor verschillende koraalsoorten, die voor praktische doeleinden kunnen worden gebruikt. Met een waterstroommeter (bijv. Swoffer stroommeters) kunnen stroomsnelheden in elk aquarium worden geoptimaliseerd, voor elke soort waarvoor gegevens beschikbaar zijn. Houd er echter rekening mee dat de onderstaande gegevens mogelijk niet van toepassing zijn op elk individu binnen een bepaalde soort, omdat bekend is dat verschillende genotypen zich anders kunnen gedragen (Osinga et al. 2011).
Om het belang van de stroomsnelheid te benadrukken, zou ik onze case study van het koraal Galaxea fascicularis willen gebruiken (Wijgerde et al. 2013). Voor poliepen in kolonies, resulteerde een waterstroomsnelheid van 10 cm s -1 in de hoogste voedingssnelheden en dus de opname van organische koolstof. Met behulp van eerder verkregen gegevens van ons laboratorium werd een nutriëntenbudget voor deze soort berekend onder verschillende stroomregimes. Hiertoe werd de input versus output van organische koolstof vergeleken. De input bestond uit koolstof geproduceerd door fotosynthese en koolstof gewonnen door voeding. De output was gebaseerd op ademhaling (dit wil zeggen de energie die wordt verbruikt door het niet-voedende, rustende dier) en uitscheiding van organisch afval. Door output af te trekken van input, werd een waarde verkregen die bekend staat als ruimte voor groei. Ruimte voor groei werd hier gedefinieerd als de koolstof die beschikbaar is voor groei, nadat is voldaan aan ademhaling en uitscheiding. Tabel 2 toont de groeimogelijkheden voor G. fascicularis bij verschillende stroomomstandigheden, en laat zien dat zeer lage en hoge stroomsnelheden resulteren in negatieve waarden. Dit suggereert dat dit koraal onder deze omstandigheden niet in staat is om voldoende prooien te vangen om weefselgroei te behouden. Hoewel deze analyse op verschillende veronderstellingen is gebaseerd, stelt het de aquariumliefhebber in staat om geschikte kweekomstandigheden te selecteren.
Meer stroming voor grotere koralen
Naast de waterstroom is de koraalgrootte van belang. De vertakte structuur van veel koralen vermindert de hoeveelheid waterstroom en licht die de binnen- en onderkant van kolonies bereikt, een fenomeen dat bekend staat als zelfschaduw. Daarom hebben grotere koralen meer licht en waterstroom nodig om de fotosynthesesnelheid en gasuitwisseling te behouden.
De voedingssnelheid per poliep (dit wil zeggen massaspecifiek) kan ook afnemen wanneer koralen groter worden, zoals gevonden voor G. fascicularis. Al deze verschijnselen verklaren gedeeltelijk waarom grotere koralen over het algemeen een lagere relatieve groeisnelheid lieten zien (hoewel de absolute groeisnelheid toeneemt omdat er meer oppervlakte is om te groeien). Dit betekent dat de aquariumliefhebber de groei van grotere koralen kan optimaliseren door te compenseren voor hun grootte, zodat dicht vertakte koralen voldoende waterstroom krijgen voor voeding en gasuitwisseling. Het in stand houden van kleine kolonies kan ook een strategie zijn om een hoge groei in de aquacultuur te behouden.
Hoge dichtheid batchvoeding of continue dosering?
Omdat voeding op lange termijn positieve effecten kan hebben op koralen, is het nuttig om ze dagelijks te voeren.
Een voordeel van batchvoeding is dat koralen zich snel kunnen voeden, zonder dat al het voer verloren gaat aan de filtratiesystemen (zie ook hieronder). Een moeilijkheid bij het voeren is echter het bepalen van de juiste dosering. Dit is afhankelijk van de biomassa die in het aquarium aanwezig is. Voor meer koralen is meer voeding nodig, onafhankelijk van het aquariumvolume. De aanwezige koraal- en andere ongewervelde dieren zullen ook van invloed zijn op de benodigde hoeveelheid voer, aangezien sommige soorten meer plankton nodig hebben dan andere. Een goede strategie is om te beginnen met een basisdosering van 100 tot 1.000 prooien per liter water en de koralen regelmatig te inspecteren. Verlies van poliepen, gebrek aan kolonie- of tentakeluitbreiding, afname van de koloniegrootte en necrose kunnen allemaal tekenen zijn van uithongering. Het doseren van aminozuren vóór een batchvoeding kan ook poliepexpansie en predatie stimuleren, mogelijk als gevolg van moleculaire receptoren die aanwezig zijn op het oppervlak van het poliep. Bij batchvoeding kan tijdelijke remming van koraalgroei worden voorkomen door overdag te voeren (zie hierboven).
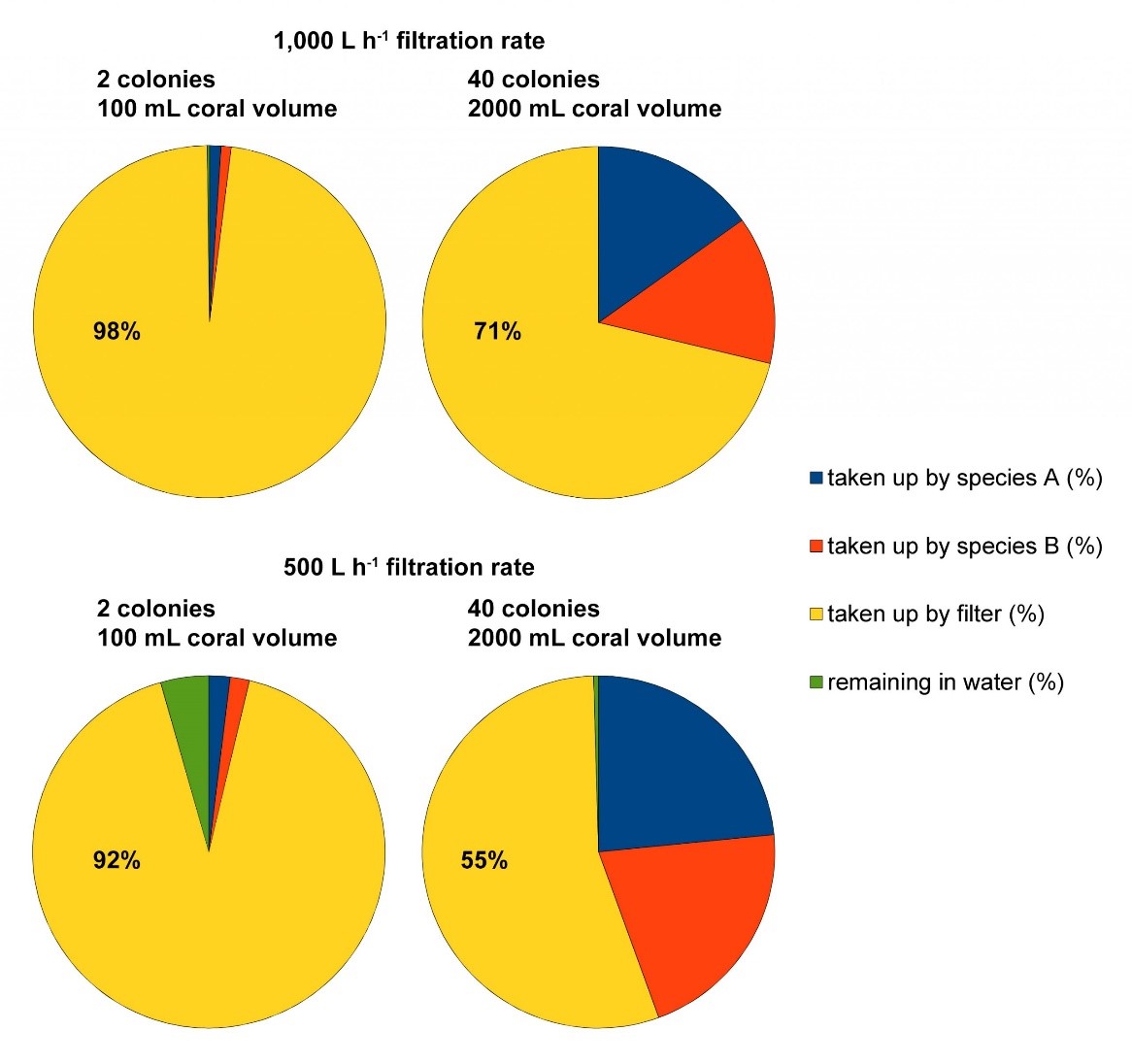
Voorbeeld van vier voedingsscenario’s berekend met de voedselsimulator (ontwikkeld door Dr. Michael Kuecken, Technische Universiteit van Dresden, en Dr. Ronald Osinga, Wageningen Universiteit), gebaseerd op een aquariumgrootte van 1.000L, 100/2.000 ml koraalvolume met een filtratie snelheid van respectievelijk 0,213 en 0,191 L h-1 ml koraal-1 voor Seriatopora hystrix (soort A) en S. caliendrum (soort B), een aquariumfiltratiesnelheid van 500/1.000 L h-1 en een voedertijd van 6 uur. Deze scenario’s laten zien hoeveel fijnstofvoedsel er in het filtersysteem terecht kan komen, vooral wanneer het koraalvolume/biomassa in het aquarium laag is.
Een andere strategie is natuurlijk plankton concentraties te handhaven (in het traject van 10 8 -10 9 algen / microbecellen en 1-10 zooplankters per liter), door langzaam pompen gekoelde culturen tot het systeem met peristaltische pompen. Voertimers kunnen ook regelmatig droogvoer geven. Deze strategie lijkt ideaal voor het behoud van azooxanthella koralen zoals Tubastraea en Dendronephthya spp.

Door culturen te doseren in een systeem met peristaltische pompen, kunnen natuurlijke planktonconcentraties worden gehandhaafd.
Controle over platwormpopulaties
Omdat acoelomorfe platwormen een nadelige invloed kunnen hebben op koralen, is het verstandig om platwormpopulaties onder controle te houden door natuurlijke vijanden in het aquarium te introduceren. Er zijn aanwijzingen dat bepaalde lipvissen (bijv. Halichoerus spp.), draken (bijv. Synchiropus splendidus ) en naaktslakken (Chelidonura varians) actief op platwormen jagen (Carl 2008; Nosratpour 2008). Chemische behandeling van koralen met anthelmintica zoals levamisol werkt goed (voor acoelomorfen), maar dit is omslachtig en kan de koraalgezondheid op lange termijn negatief beïnvloeden.

Platwormen, hier gehost door Goniopora spp. (links en rechts) en Euphyllia paraancora (midden), kunnen met natuurlijke vijanden onder controle worden gehouden.
Gebruik van planktonbesparende filtersystemen
Tot nu toe heb ik de impact van aquariumfiltratiesystemen op koraalvoeding niet besproken. Aangezien de meeste aquaria zijn uitgerust met schuimfractionators (eiwitafschuimers), met het vermogen om kleine deeltjes te verwijderen, is het logisch dat deze filters enige invloed zullen hebben op de beschikbaarheid van voedsel voor koralen. Inderdaad, wanneer droog visvoer of fytoplanktonculturen aan een aquarium worden toegevoegd, komt een deel hiervan al snel in de opvangbeker van de skimmer terecht. Dit is gemakkelijk te zien wanneer het voer of de cultuur een schijnbare kleur heeft, zoals oranje of groen.
De voor de hand liggende vragen die hieruit voortvloeien zijn; hoeveel voedseldeeltjes komen er eigenlijk in het filtersysteem terecht, en hoeveel voedsel zal worden gegeten door de koralen en andere filtervoedende organismen? Hiertoe werd in het kader van het onderzoeksproject CORALZOO een voedselsimulator ontwikkeld . Met behulp van schattingen van de opnamesnelheid van koraalvoedsel, gebaseerd op laboratoriumexperimenten (Wijgerde en Osinga 2007, niet-gepubliceerde gegevens), en informatie zoals koraalvolume (een maat voor koraalbiomassa), prooiconcentratie, systeemwatervolume en aquariumfiltratiesnelheden, kan het lot van voedseldeeltjes die aan het systeem worden toegevoegd, worden berekend.

Koraalvoeding is van vitaal belang voor een succesvolle koraalaquacultuur.
Om te illustreren hoe nuttig dit programma is, heb ik twee scenario’s berekend. In de eerste twee scenario’s is een aquarium van 1000 liter gevuld met ofwel twee (een Seriatopora hystrix en een S. caliendrum kolonie) of veertig (twintig S. hystrix en twintig S. caliendrum kolonies) koralen van gemiddelde grootte (50 ml volume per kolonie). ). De aquariumfiltratiesnelheid is ingesteld op 1.000 L h -1 , gebaseerd op een netto wateromzet tussen het aquarium en de opvangbak van 1.000 L h -1 , en een eiwitafschuimer uitgerust met een 1.000 L h -1 pomp. Dus de verblijftijd van de voedseldeeltjes (Artemia-nauplii) in het aquarium is precies een uur. In de twee twee scenario’s zijn de bezettingsdichtheden gelijk, maar wordt de filtratiesnelheid gehalveerd tot 500 L h -1 . Dus de verblijftijd van de voedseldeeltjes (Artemia-nauplii) is gestegen tot twee uur. Na zes uur zijn alle scenario’s bijna in evenwicht en worden de volgende datasets verkregen; in de eerste twee scenario’s is 98% en 71% van het voedsel door het filter geconsumeerd. In de tweede twee zijn deze waarden 92% en 55%. Hoewel dit theoretische voorbeeld een aantal beperkingen kent, zoals de aanname dat alle deeltjes door de eiwitafschuimer worden verwijderd, laat het wel duidelijk zien dat mechanische filters (waaronder biofilters en zandfilters) leiden tot een aanzienlijke verspilling van voedsel. Dit afval kan worden verminderd door de bezettingsdichtheid van koralen te vergroten en/of door de filtercapaciteit van het aquarium (continu of tijdelijk) te verminderen, zodat de koralen zich meer kunnen voederen. Aangezien filtratie vereist is om een hoge waterkwaliteit te behouden, is het duidelijk dat deze wisselwerking tussen voedselbeschikbaarheid en waterkwaliteit kan worden omzeild door planktonbesparende filtratiesystemen te gebruiken, waaronder denitrificerende zandbedden zoals Dynamic Mineral Control (DyMiCo) en algengraswassers (Wijgerde 2012a,b). Omdat de nevenvoordelen van de eiwitafschuimer verloren gaan (dit wil zeggen het behoud van een hoge zuurstofverzadiging en pH via beluchting van het water), is het belangrijk om dit te compenseren, vooral ‘s nachts. Dit kan door het water te beluchten met luchtpompen, of door de bezettingsdichtheid van koralen en vissen te beperken.
Slotopmerkingen
Alles bij elkaar genomen, is het duidelijk dat koralen zich op vele manieren voeden, waardoor ze echte alleseters zijn. Deze kennis kan effectief worden gebruikt door aquariumliefhebbers en zo bijdragen aan koraalaquacultuur en een succesvolle aquariumhobby. Door koralen te voorzien van een verscheidenheid aan voeding, naast voldoende licht, geschikte waterbeweging en schoon water, zullen ze groeien en gedijen in het aquarium. Toekomstig onderzoek zal ongetwijfeld nieuwe wegen blootleggen waarlangs deze fascinerende dieren zich voeden, waardoor aquariumliefhebbers hun beschikbare kweekmethoden verder kunnen verfijnen.
Referenties
Allemand D, Ferrier-Pagès C, Furla P, Houlbrèque F, Puverel S, Reynaud S, Tambutté É, Tambutté S, Zoccola D (2004) Biomineralisatie in rifbouwende koralen: van moleculaire mechanismen tot milieucontrole. CR Palevol 3:453-467
Allemand D, Tambutté E, Girard JP, Jaubert J (1998) Organische matrixsynthese in het scleractinian koraal Stylophora pistillata: rol in biomineralisatie en mogelijk doelwit van het organotin tribulyltine. J Exp Biol 201:2001-2009
Anthony KRN (1999) Koraalsuspensie die zich voedt met fijne deeltjes. J Exp maart Biol Ecol 232:85-106
Anthony KRN (2000) Verbeterde capaciteit voor het voeden van deeltjes van koralen op troebele riffen (Great Barrier Reef, Australië). Koraalriffen 19:59-67
Anthony KRN, Fabricius K (2000) Verschuivende rollen van heterotrofie en autotrofie in koraaldynamica onder variërende troebelheid. J Exp Mar Biol Ecol 252:221-253
Barneah O, Brickner I, Hooge M, Weis VM, LaJeunesse TC, Benayahu Y (2007) Driepartijensymbiose: acoelomorfe wormen, koralen en eencellige algensymbionten in Eilat (Rode Zee). Mar Biol 151: 1215-1223
Bo M (2009) Antipatharians - deel twee: ecologie. Coralscience.org, www.coralscience.org
Carl M (2008) Roofdieren en plagen van koralen in gevangenschap, 31-36. In: Leewis RJ, Janse M (Eds) Advances in Coral Husbandry in Public Aquaria - Public Aquarium Husbandry Series, Volume 2, Burgers’ Zoo, Arnhem, Nederland. 444 p
Dai CF, Lin MC (1993) De effecten van stroming op het voeden van drie gorgonen uit het zuiden van Taiwan. J Exp Mar Biol Ecol 173:57-69
Davies AJ, Duineveld GCA, Lavaleye MSS, Bergman MJN, Van Haren H et al. (2009) Downwelling en diepwaterbodemstromingen als voedselvoorzieningsmechanismen voor het koudwaterkoraal Lophelia pertusa (Scleractinia) in het Mingulay Reef Complex. Limnol Oceanogr 54:620-629
Elyakova LA, Shevchenko NM, Avaeva SM (1981) Een vergelijkende studie van carbohydrase-activiteiten bij ongewervelde zeedieren, Comp. Biochem Fysiol 69B:905-908
Erftemeijer PLA, Riegl B, Hoeksema BW, Todd PA (2012) Milieueffecten van baggeren en andere sedimentverstoringen op koralen: een overzicht. Mar Poll Bull 64: 1737-1765
Fabricius KE, Alderslade P (2001) Zachte koralen en zeewaaiers - Een uitgebreide gids voor de tropische ondiepe watersoorten van de Centraal-West Pacific, de Indische Oceaan en de Rode Zee. Australian Institute of Marine Science, Townsville, Australië. 264 euro
Fabricius KE, Benayahu Y, Genin A (1995b) Herbivoor in asymbiotische zachte koralen. Wetenschap 268:90-92
Fabricius KE, Genin A, Benayahu Y (1995a) Stroomafhankelijke herbivoor en groei in zooxanthellae-vrije zachte koralen. Limnol Oceanogr 40: 1290-1301
Ferrier-Pagès C, Allemand D, Gattuso JP, Jaubert J, Rassoulzadegan F (1998a) Microheterotrofie in het zoöxanthellaatkoraal Stylophora pistillata : effecten van licht en ciliaatdichtheid. Limnol Oceanogr 43:1639-1648
Ferrier-Pagès C, Gattuso JP, Cauwet G, Jaubert J, Allemand D (1998b) Afgifte van opgeloste organische koolstof en stikstof door het zoöxanthellaatkoraal Galaxea fascicularis. Mar Ecol Prog Ser 172:265-274
Ferrier-Pagès C, Hoogenboom M, Houlbrèque F (2011) De rol van plankton in koraaltrofodynamica, 215-229. In: Dubinsky Z, Stambler N (Eds), Koraalriffen: een ecosysteem in transitie. Springer, Dordrecht, Nederland
Ferrier-Pagès C, Witting J, Tambutté E, Sebens KP (2003) Effect van natuurlijke zoöplanktonvoeding op de weefsel- en skeletgroei van het scleractinian koraal Stylophora pistillata. Koraalriffen 22:229-240
Fine M, Loya Y (2002) Endolithische algen: een alternatieve bron van foto-assimilaten tijdens het bleken van koraal. Proc R Soc B 269: 1205-1210
Goreau TF, Goreau NI, Yonge CM (1971) Rifkoralen: autotrofen of heterotrofen? Biologisch Bulletin 141:247-260
Grover R, Maguer JF, Allemand D, Ferrier-Pagès C (2006) Ureumopname door het scleractinian koraal Stylophora pistillata. J Exp maart Biol Ecol 332:216-225
Grover R, Maguer JF, Allemand D, Ferrier-Pagès C (2008) Opname van opgeloste vrije aminozuren (DFAA) door het scleractinian koraal Stylophora pistillata. J Exp Biol 211:860-865
Helmuth B, Sebens K (1993) De invloed van koloniemorfologie en oriëntatie om te stromen bij het vangen van deeltjes door het scleractinian koraal Agaricia agaricites (Linnaeus). J Exp Mar Biol Ecol 165:251-278
Hoeksema BW, Farenzena ZT (2012) Weefselverlies bij koralen aangetast door acoelomorph platwormen ( Waminoa sp.). Koraalriffen 31:869
Houlbrèque F, Ferrier-Pagès C (2009) Heterotrofie in tropische scleractinian koralen. Biol Rev Camb Philos 84:1-17
Houlbrèque F, Tambutté E, Richard C, Ferrier-Pagès C (2004) Het belang van een microdieet voor scleractinian koralen. Mar Ecol Prog Ser 282:151-160
Hunter T (1989) Suspensievoeding in oscillerende stroming: het effect van koloniemorfologie en stromingsregime op planktonvangst door de hydroïde Obelia longissima. Biol Bull 176:41-49
Jimenez IM, Kühl M, Larkum AWD, Ralph PJ (2011) Effecten van stroming en morfologie op de thermische grenslaag van koralen. JR Soc-interface 8:1785-1795
Lai S, Gillis LG, Mueller C, Bouma TJ, Guest JR, Last KS, Ziegler AD, Todd PA (2013) Eerste experimentele bewijs van koralen die zich voeden met zeegras. Koraalriffen DOI 10.1007/s00338-013-1062-9
Leal MC, Ferrier-Pagès C, Calado R, Thompson ME, Frischer ME, Nejstgaard JC (2013) Koraalvoeding met microalgen beoordeeld met moleculaire trofische markers. Mol Ecol doi: 10.1111/mec.12486
Lesser MP, Falcón LI, Rodríguez-Román A, Enríquez S, Hoegh-Guldberg O, Iglesias-Prieto R (2007) Stikstoffixatie door symbiotische cyanobacteriën vormt een bron van stikstof voor het scleractinische koraal Montastraea cavernosa. Mar Ecol Prog Ser 346:143-152
Lewis JB (2006) Biologie en ecologie van de hydrocoral Millepora op koraalriffen. Adv Mar Biol 50:1-55
Lin MC, Liao CM, Dai CF (2002) Modellering van de effecten van verzadiging op de voedingssnelheid van een koloniale suspensievoeder, Acanthogorgia vegae, in een circulerend systeem onder laboratoriumomstandigheden. Zool Stud 41:355-365
Muscatine L (1990) De rol van symbiotische algen in koolstof- en energieflux in rifkoralen, 755-87. In: Dubinsky Z (Ed), Koraalriffen: ecosystemen van de wereld 25. Elsevier, Amsterdam, Nederland
Naumann MS, Mayr C, Struck U, Wild C (2010) Koraalslijmstabiele isotoopsamenstelling en labeling: experimenteel bewijs voor slijmopname door epizoïsche acoelomorfe wormen. Mar Biol 157:2521-2531
Nosratpour F (2008) Waarnemingen van een polyclad platworm die acroporide koralen in gevangenschap aantast. In: Leewis RJ, Janse M (Eds) Vooruitgang in koraalhouderij in openbare aquaria - Public Aquarium Husbandry Series, Volume 2, Burgers’ Zoo, Arnhem, 37-46
Osinga R, Schutter M, Griffioen B, Wijffels RH, Verreth JAJ, Shafir S, Henard S, Taruffi M, Gili C, Lavorano S (2011) De biologie en economie van koraalgroei. Mar Biotechnol 13:658-671
Osinga R, Schutter M, Wijgerde T, Rinkevich B, Shafir S, Shpigel M, Luna GM, Danovaro R, Bongiorni L, Deutsch A, Kuecken M, Hiddinga B, Janse M, McLeod A, Gili C, Lavorano S, Henard S , Barthelemy D, Westhoff G, Baylina N, Santos E, Weissenbacher A, Kuba M, Jones R, Leewis R, Petersen D, Laterveer M (2012) Het CORALZOO-project: een samenvatting van vier jaar openbare aquariumwetenschap. Journal Mar Biol Assoc UK 92:753-768
Picciano M, Ferrier-Pagès C (2007) Inname van pico- en nanoplankton door het mediterrane rode koraal Corallium rubrum. Mar Biol 150:773-782
Ribes M, Coma R, Gili JM (1999) Heterogene voeding in benthische suspensievoeders: het natuurlijke dieet en de begrazingssnelheid van de gematigde gorgonian Paramuricea clavata (Cnidaria: Octocorallia) gedurende een jaarcyclus. Mar Ecol Prog Ser 183:125-137
Roff G, Dove SG, Dunn SR (2009) Mesenteriale filamenten maken een schone zwaai van substraten voor koraalgroei. Koraalriffen 28:79
Schutter M, Crocker J, Paijmans A, Janse M, Osinga R, Verreth AJ, Wijffels RH (2010) Het effect van verschillende stroomregimes op de groei en stofwisseling van het scleractinian koraal Galaxea fascicularis. Koraalriffen 29:737-748
Schutter M, Kranenbarg S, Wijffels RH, Verreth JAJ, Osinga R (2011) Modificatie van lichtgebruik voor skeletgroei door waterstroom in het scleractinian koraal Galaxea fascicularis. Mar Biol 158:769-777
Sebens KP, Grace SP, Helmuth B, Maney Jr EJ, Miles JS (1998) Waterstroom en prooivangst door drie scleractinian koralen, Madracis mirabilis, Montastrea cavernosa en Porites porites, in een veldbehuizing. Maart Biol 131: 347-360
Swanson R, Hoegh-Guldberg O (1998) Aminozuursynthese in de symbiotische zeeanemoon Aiptasia pulchella. Mar Biol 131:83-93
Tsounis G, Rossi S, Laudien J, Bramanti L, Fernández N, Gili JM, Arntz W (2006) Dieet- en seizoensgebonden prooi-vangstpercentages in het mediterrane rode koraal (Corallium rubrum L.). Mar Biol 149:313-325
Wijgerde T (2012a) Verbeterde houderij van ongewervelde zeedieren met behulp van een innovatieve filtratietechnologie – deel één: DyMiCo. Gevorderde aquariumliefhebber 11(2)
Wijgerde T (2012b) Verbeterde houderij van ongewervelde zeedieren met behulp van een innovatieve filtratietechnologie – deel twee: resultaten met twee 12 m 3 DyMiCo-systemen. Gevorderde aquariumliefhebber 11 (3)
Wijgerde T (2013a) Zooxanthellae: biologie en isolatie voor wetenschappelijk onderzoek. Gevorderde aquariumliefhebber 12(5)
Wijgerde T (2013b) Heterotrofe voeding, groei en nutriëntenbudget in het scleractinian koraal Galaxea fascicularis. Proefschrift, Wageningen Universiteit, Wageningen, Nederland
Wijgerde T, Diantari R, Lewaru MW, Verreth JAJ, Osinga R (2011a) Extracoelenterische zoöplanktonvoeding is een belangrijk mechanisme voor het verwerven van voedingsstoffen voor het scleractinian koraal Galaxea fascicularis. J Exp Biol 214: 3351-3357
Wijgerde T, Henkemans P, Osinga R (2012a) Effecten van bestraling en lichtspectrum op de groei van het scleractinian koraal Galaxea fascicularis - Toepasbaarheid van LEP- en LED-verlichting op koraalaquacultuur. Aquacultuur 344-349: 188-193
Wijgerde T, Jurriaans S, Hoofd M, Verreth JAJ, Osinga R (2012b) Zuurstof en heterotrofie beïnvloeden de verkalking van het scleractinian koraal Galaxea fascicularis. PLoS ONE 7 (12): e52702. doi:10.1371/journal.pone.0052702
Wijgerde T, Schots P, van Onselen E, Janse M, Karruppannan E, Verreth JAJ, Osinga R (2012c) Epizoïsche acoelomorfe platwormen verminderen de voeding van zoöplankton door het scleractinian koraal Galaxea fascicularis. Biol Open 2:10-17
Wijgerde T, Spijkers P, Karruppannan E, Verreth JAJ, Osinga R (2012d) Waterstroom beïnvloedt de voeding van zoöplankton door het scleractinian koraal Galaxea fascicularis op poliep- en kolonieniveau . J Mar Biol doi: 10.1155/2012/854849
Wijgerde T, Spijkers P, Verreth J, Osinga R (2011b) Epizoïsche acoelomorfe platwormen concurreren met hun koraalgastheer voor zoöplankton. Koraalriffen 30:665
Purser A, Larsson AI, Thomsen L, van Oevelen D (2010) De invloed van stroomsnelheid en voedselconcentratie op zoöplankton-opnamesnelheden van Lophelia pertusa (Scleractinia). J Exp maart Biol Ecol 395:55–62
Inloggen
Registreren
Foto van de maand

Blauwe mandarijnpitvis Synchiropus splendidus
Foto: Sander Hemmes